

Dopamine D1-like receptors modulate synchronized oscillations in the hippocampal-prefrontal-amygdala circuit in contextual fear
- PMID: 37848657
- PMCID: PMC10582086
- DOI: 10.1038/s41598-023-44772-6
Dopamine D1-like receptors modulate synchronized oscillations in the hippocampal-prefrontal-amygdala circuit in contextual fear
Abstract
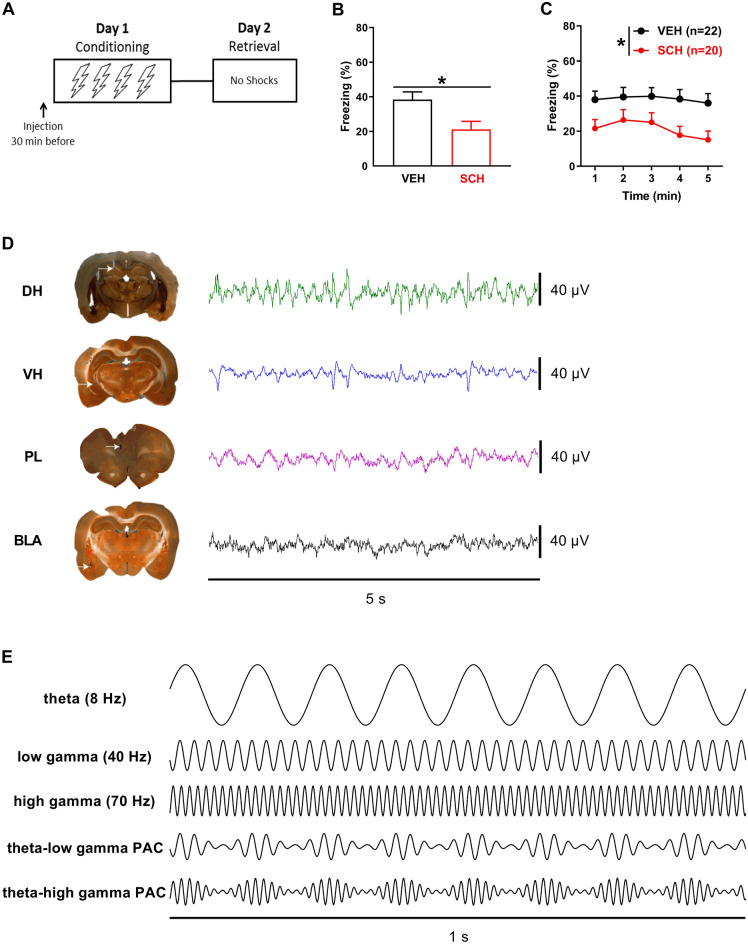
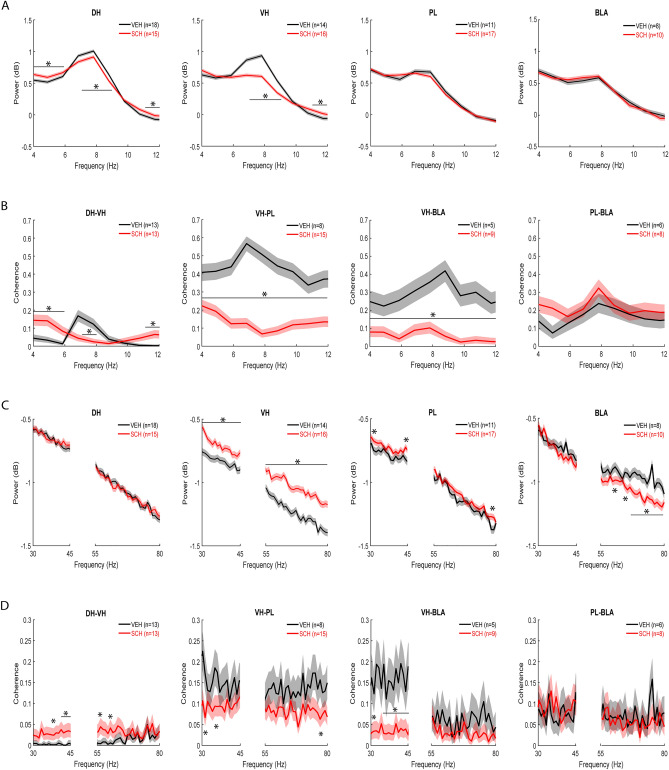
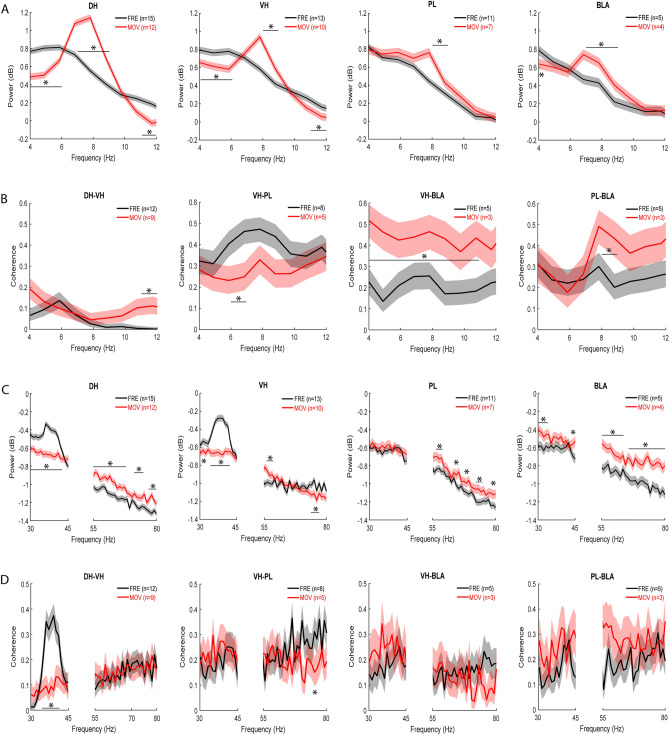
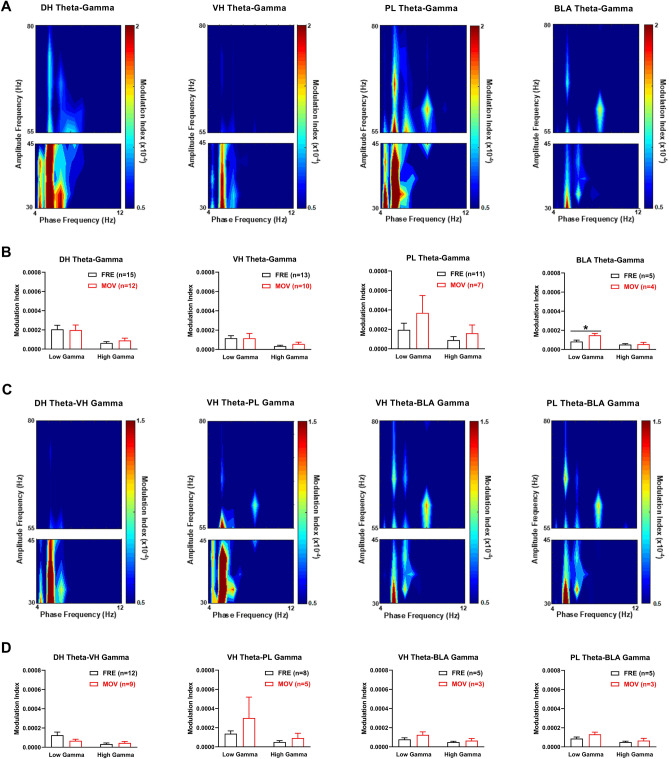
Contextual fear conditioning (CFC) is mediated by a neural circuit that includes the hippocampus, prefrontal cortex, and amygdala, but the neurophysiological mechanisms underlying the regulation of CFC by neuromodulators remain unclear. Dopamine D1-like receptors (D1Rs) in this circuit regulate CFC and local synaptic plasticity, which is facilitated by synchronized oscillations between these areas. In rats, we determined the effects of systemic D1R blockade on CFC and oscillatory synchrony between dorsal hippocampus (DH), prelimbic (PL) cortex, basolateral amygdala (BLA), and ventral hippocampus (VH), which sends hippocampal projections to PL and BLA. D1R blockade altered DH-VH and reduced VH-PL and VH-BLA synchrony during CFC, as inferred from theta and gamma coherence and theta-gamma coupling. D1R blockade also impaired CFC, as indicated by decreased freezing at retrieval, which was characterized by altered DH-VH and reduced VH-PL, VH-BLA, and PL-BLA synchrony. This reduction in VH-PL-BLA synchrony was not fully accounted for by non-specific locomotor effects, as revealed by comparing between epochs of movement and freezing in the controls. These results suggest that D1Rs regulate CFC by modulating synchronized oscillations within the hippocampus-prefrontal-amygdala circuit. They also add to growing evidence indicating that this circuit synchrony at retrieval reflects a neural signature of learned fear.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
