

Flight muscle size reductions and functional changes following long-distance flight under variable humidity conditions in a migratory warbler
- PMID: 37849053
- PMCID: PMC10582281
- DOI: 10.14814/phy2.15842
Flight muscle size reductions and functional changes following long-distance flight under variable humidity conditions in a migratory warbler
Abstract
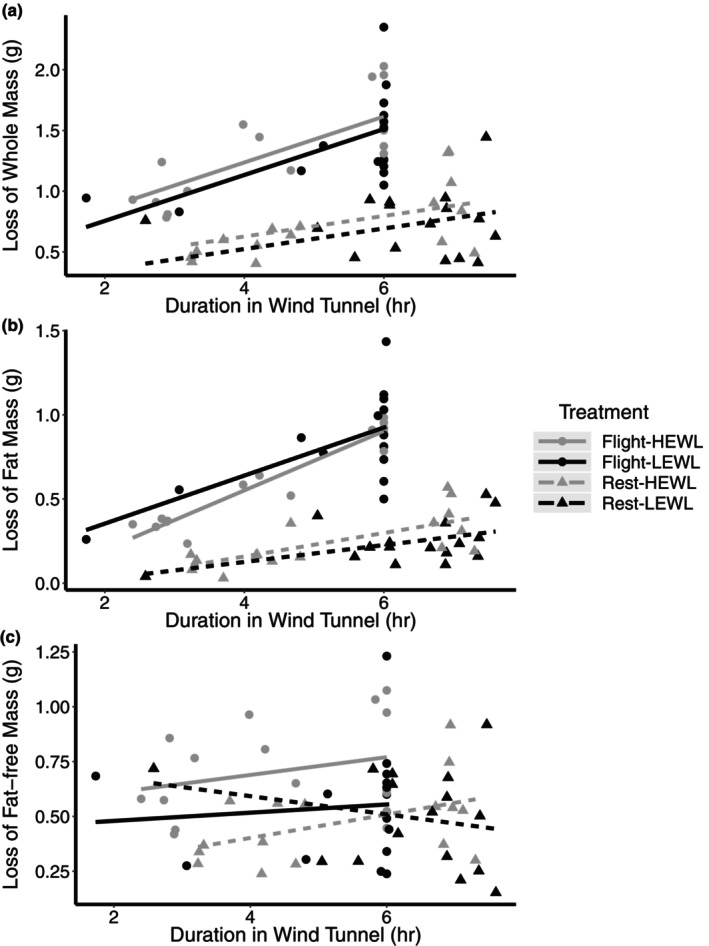
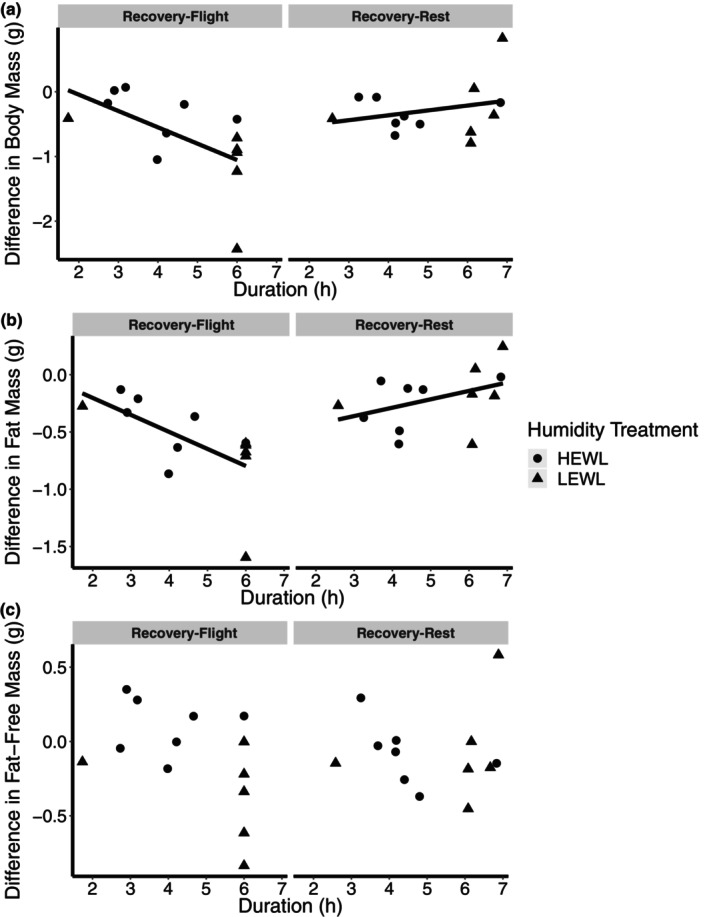
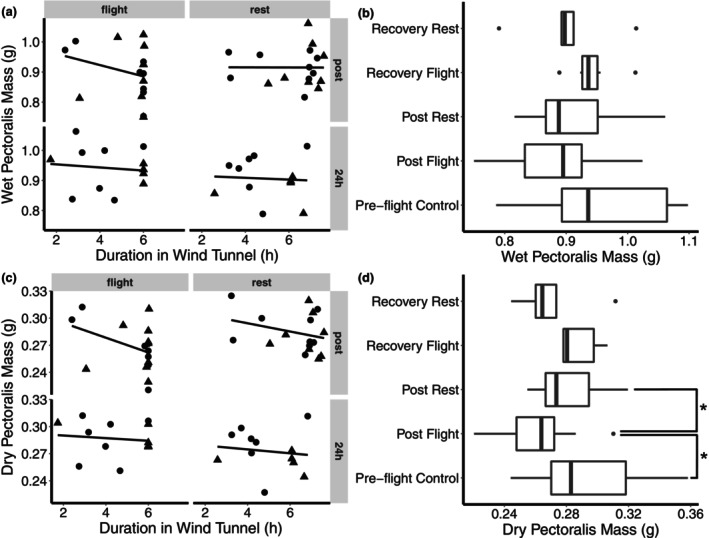
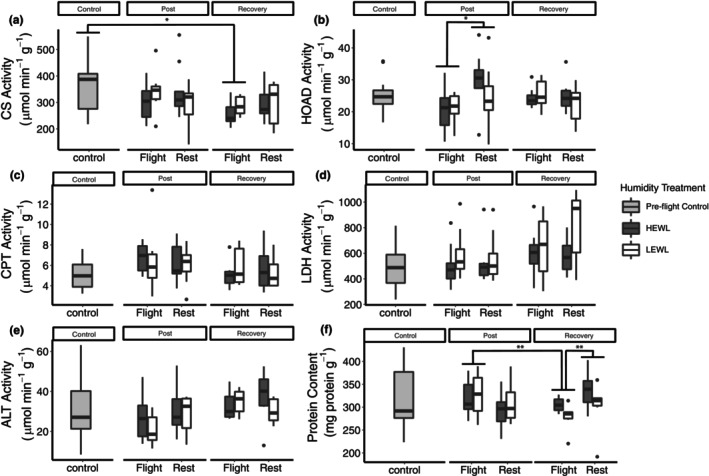
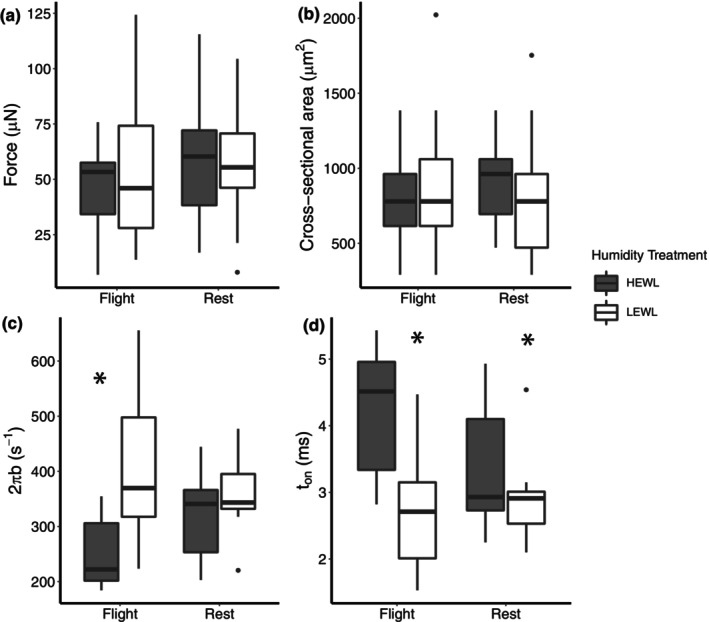
Bird flight muscle can lose as much as 20% of its mass during a migratory flight due to protein catabolism, and catabolism can be further exacerbated under dehydrating conditions. However, the functional consequences of exercise and environment induced protein catabolism on muscle has not been examined. We hypothesized that prolonged flight would cause a decline in muscle mass, aerobic capacity, and contractile performance. This decline would be heightened for birds placed under dehydrating environmental conditions, which typically increases lean mass losses. Yellow-rumped warblers (Setophaga coronata) were exposed to dry or humid (12 or 80% relative humidity at 18°C) conditions for up to 6 h while at rest or undergoing flight. The pectoralis muscle was sampled after flight/rest or after 24 h of recovery, and contractile properties and enzymatic capacity for aerobic metabolism was measured. There was no change in lipid catabolism or force generation of the muscle due to flight or humidity, despite reductions in pectoralis dry mass immediately post-flight. However, there was a slowing of myosin-actin crossbridge kinetics under dry compared to humid conditions. Aerobic and contractile function is largely preserved after 6 h of exercise, suggesting that migratory birds preserve energy pathways and function in the muscle.
Keywords: atrophy; dehydration; exercise; migration; skeletal muscle.
© 2023 The Authors. Physiological Reports published by Wiley Periodicals LLC on behalf of The Physiological Society and the American Physiological Society.
Conflict of interest statement
No conflicts of interest, financial, or otherwise, are declared by the authors.
Figures
References
-
- Battley, P. F. , Piersma, T. , Dietz, M. W. , Tang, S. , Dekinga, A. , & Hulsman, K. (2000). Empirical evidence for differential organ reductions during trans–oceanic bird flight. Proceedings of the Royal Society of London Series B: Biological Sciences, 267, 191–195. 10.1098/rspb.2000.0986 - DOI - PMC - PubMed
-
- Bauchinger, U. , & Biebach, H. (1998). The role of protein during migration in passerine birds. Biol Conserv Fauna, 102, 299–305.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
