

Tr1 Cells Emerge and Suppress Effector Th17 Cells in Glomerulonephritis
- PMID: 37850963
- PMCID: PMC10656435
- DOI: 10.4049/jimmunol.2300305
Tr1 Cells Emerge and Suppress Effector Th17 Cells in Glomerulonephritis
Abstract
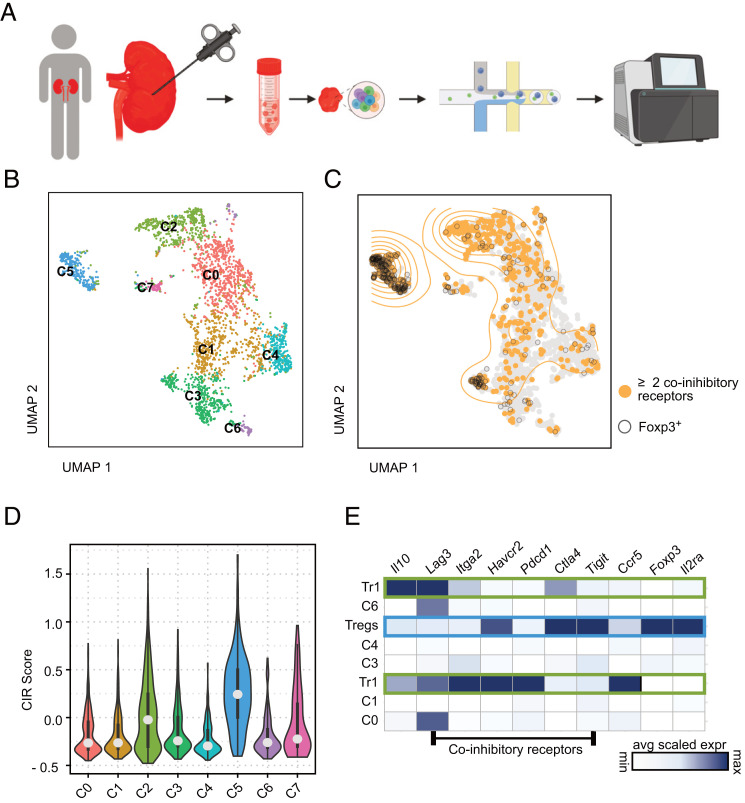
T regulatory type 1 (Tr1) cells, which are defined by their regulatory function, lack of Foxp3, and high expression of IL-10, CD49b, and LAG-3, are known to be able to suppress Th1 and Th17 in the intestine. Th1 and Th17 cells are also the main drivers of crescentic glomerulonephritis (GN), the most severe form of renal autoimmune disease. However, whether Tr1 cells emerge in renal inflammation and, moreover, whether they exhibit regulatory function during GN have not been thoroughly investigated yet. To address these questions, we used a mouse model of experimental crescentic GN and double Foxp3mRFP IL-10eGFP reporter mice. We found that Foxp3neg IL-10-producing CD4+ T cells infiltrate the kidneys during GN progression. Using single-cell RNA sequencing, we could show that these cells express the core transcriptional factors characteristic of Tr1 cells. In line with this, Tr1 cells showed a strong suppressive activity ex vivo and were protective in experimental crescentic GN in vivo. Finally, we could also identify Tr1 cells in the kidneys of patients with antineutrophil cytoplasmic autoantibody-associated GN and define their transcriptional profile. Tr1 cells are currently used in several immune-mediated inflammatory diseases, such as T-cell therapy. Thus, our study provides proof of concept for Tr1 cell-based therapies in experimental GN.
Copyright © 2023 by The American Association of Immunologists, Inc.
Conflict of interest statement
The authors have no financial conflicts of interest.
Figures






References
-
- Krebs, C. F., Turner J.-E., Paust H.-J., Kapffer S., Koyro T., Krohn S., Ufer F., Friese M. A., Flavell R. A., Stockinger B., et al. . 2016. Plasticity of Th17 cells in autoimmune kidney diseases. J. Immunol. 197: 449–457. - PubMed
-
- Tipping, P. G., Holdsworth S. R.. 2006. T cells in crescentic glomerulonephritis. J. Am. Soc. Nephrol. 17: 1253–1263. - PubMed
-
- Paust, H.-J., Turner J.-E., Riedel J.-H., Disteldorf E., Peters A., Schmidt T., Krebs C., Velden J., Mittrücker H.-W., Steinmetz O. M., et al. . 2012. Chemokines play a critical role in the cross-regulation of Th1 and Th17 immune responses in murine crescentic glomerulonephritis. Kidney Int. 82: 72–83. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
