

The ClpX protease is essential for inactivating the CI master repressor and completing prophage induction in Staphylococcus aureus
- PMID: 37852980
- PMCID: PMC10584840
- DOI: 10.1038/s41467-023-42413-0
The ClpX protease is essential for inactivating the CI master repressor and completing prophage induction in Staphylococcus aureus
Abstract
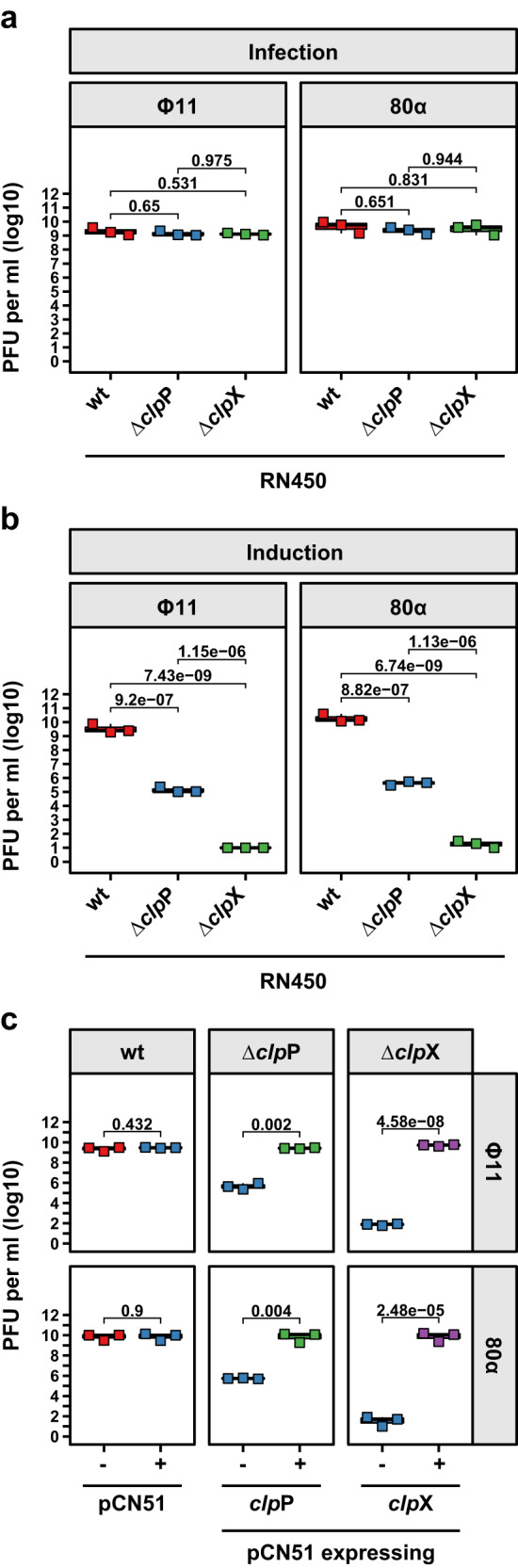
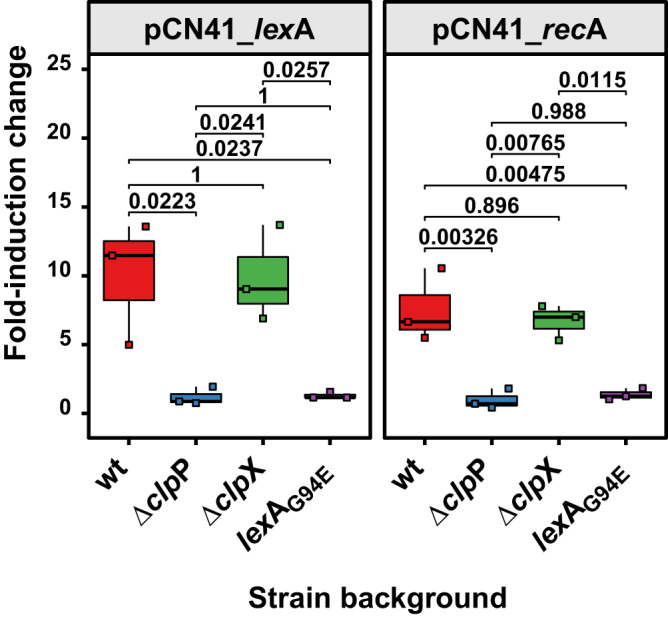
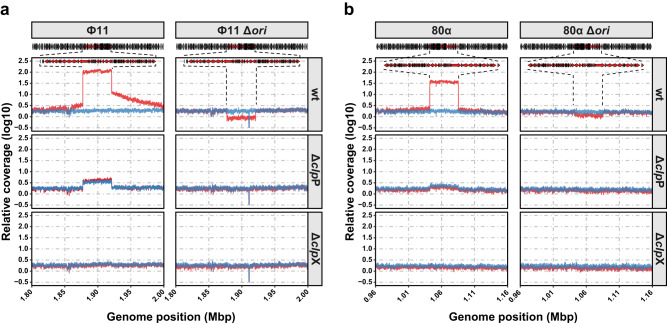
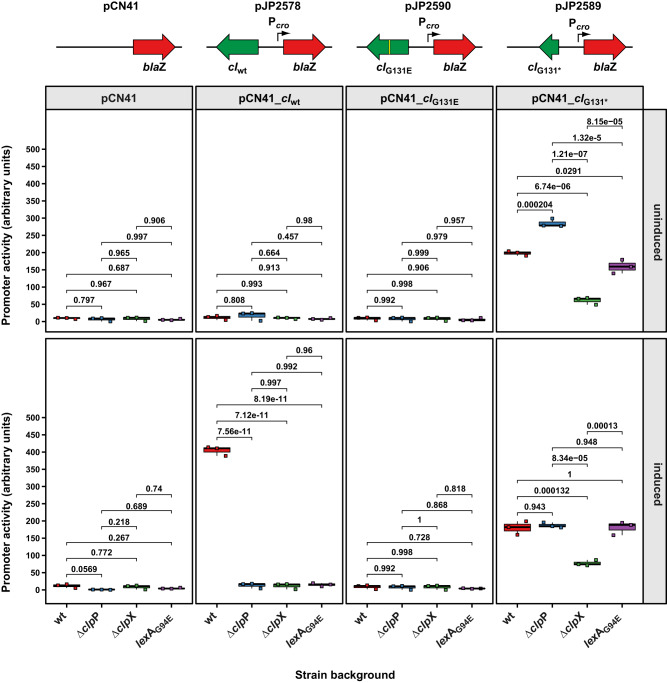
Bacteriophages (phages) are the most abundant biological entities on Earth, exerting a significant influence on the dissemination of bacterial virulence, pathogenicity, and antimicrobial resistance. Temperate phages integrate into the bacterial chromosome in a dormant state through intricate regulatory mechanisms. These mechanisms repress lytic genes while facilitating the expression of integrase and the CI master repressor. Upon bacterial SOS response activation, the CI repressor undergoes auto-cleavage, producing two fragments with the N-terminal domain (NTD) retaining significant DNA-binding ability. The process of relieving CI NTD repression, essential for prophage induction, remains unknown. Here we show a specific interaction between the ClpX protease and CI NTD repressor fragment of phages Ф11 and 80α in Staphylococcus aureus. This interaction is necessary and sufficient for prophage activation after SOS-mediated CI auto-cleavage, defining the final stage in the prophage induction cascade. Our findings unveil unexpected roles of bacterial protease ClpX in phage biology.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
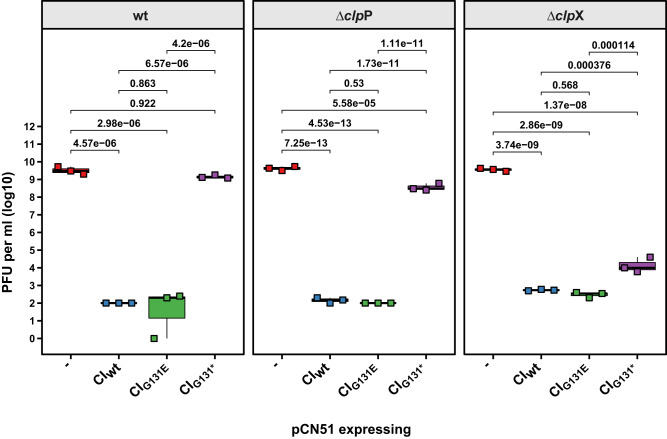

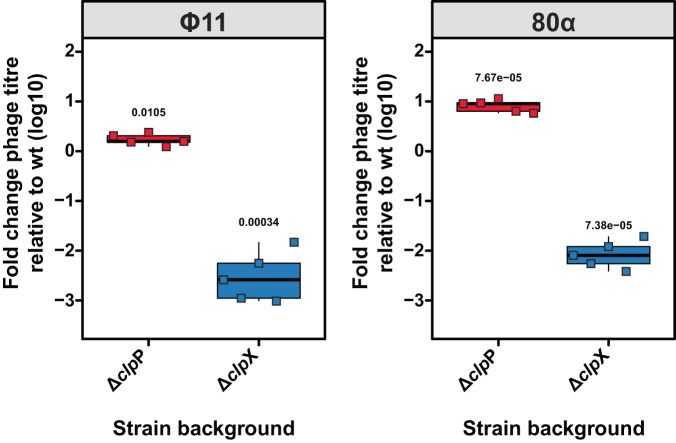
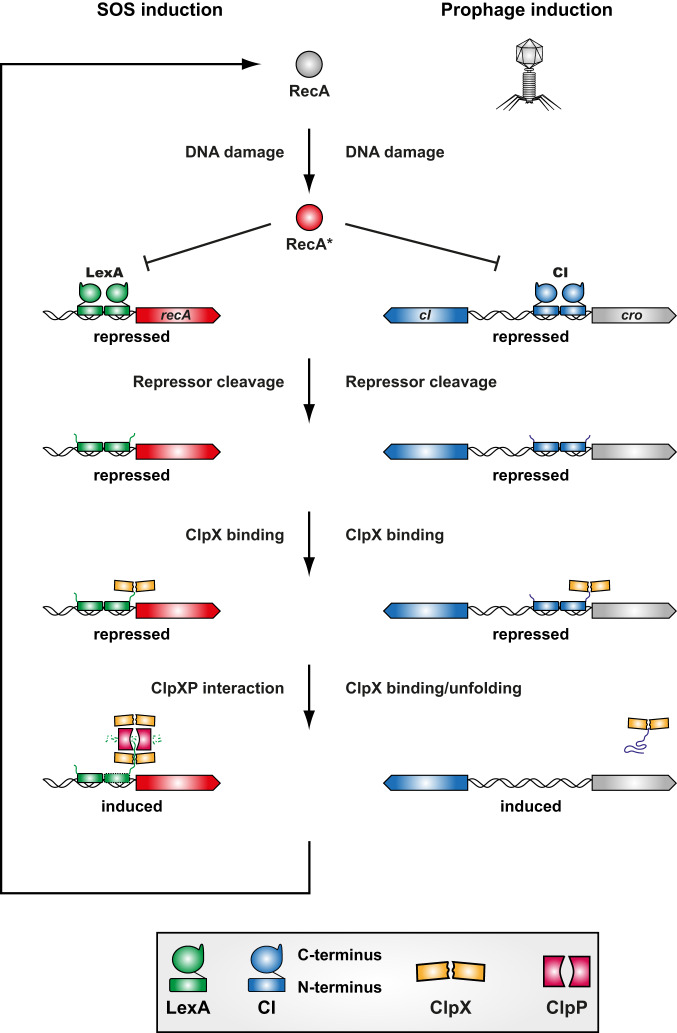
References
-
- Comeau AM, et al. Exploring the prokaryotic virosphere. Res. Microbiol. 2008;159:306–313. - PubMed
-
- Ptashne M. A genetic switch: phage λ revisited, 3rd edn. Cold Spring Harbor Laboratory Press (2004).
Publication types
MeSH terms
Substances
Grants and funding
- MR/S00940X/2/MRC_/Medical Research Council/United Kingdom
- 201531/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- MR/M003876/1 /MRC_/Medical Research Council/United Kingdom
- BB/N002873/1 /BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/S00940X/1/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
