

Macromolecular condensation buffers intracellular water potential
- PMID: 37853127
- PMCID: PMC10665201
- DOI: 10.1038/s41586-023-06626-z
Macromolecular condensation buffers intracellular water potential
Erratum in
-
Author Correction: Macromolecular condensation buffers intracellular water potential.Nature. 2024 Apr;628(8009):E4. doi: 10.1038/s41586-024-07346-8. Nature. 2024. PMID: 38589575 Free PMC article. No abstract available.
Abstract
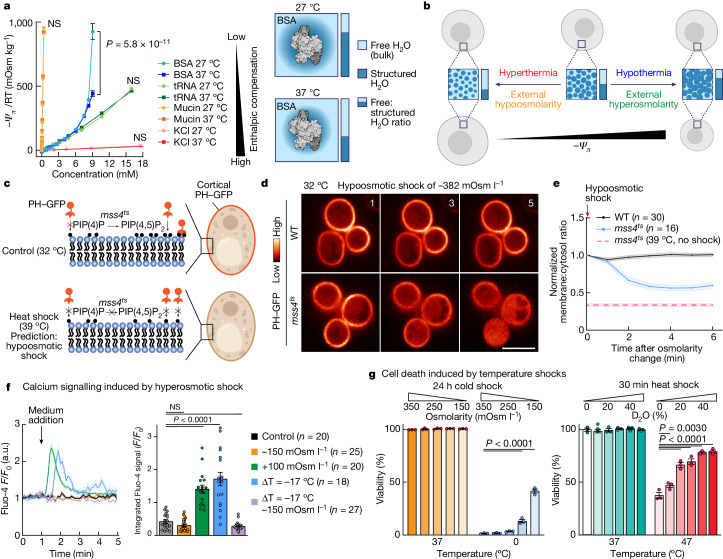
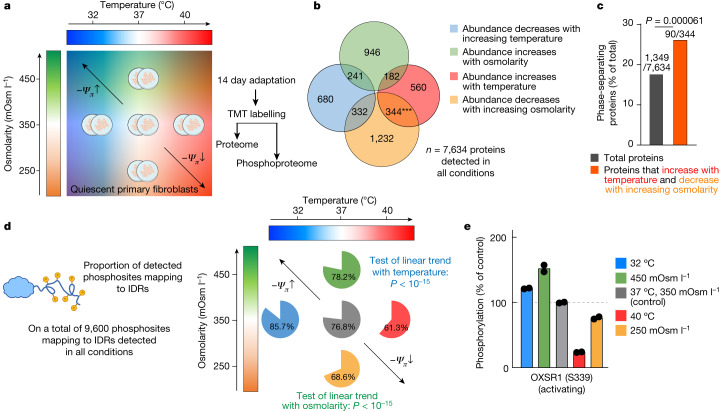
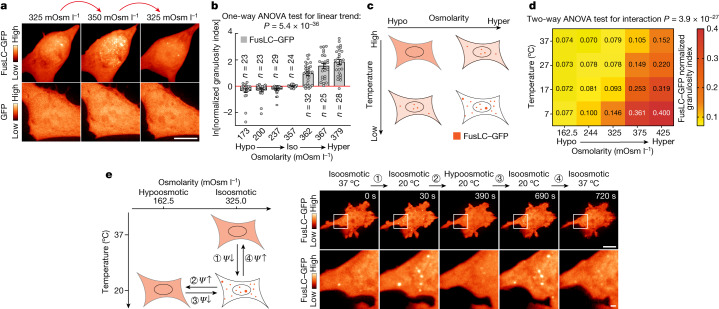
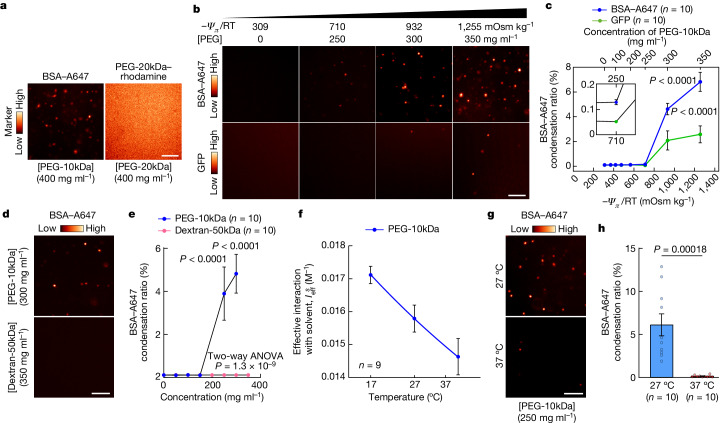
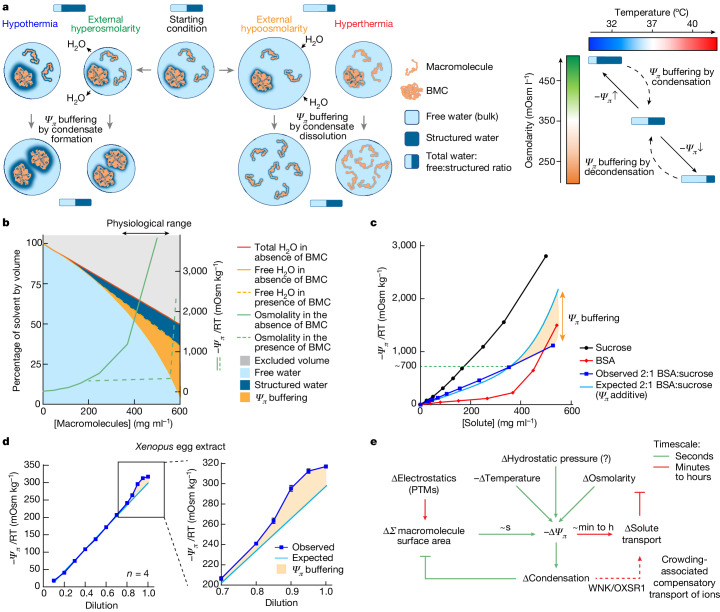
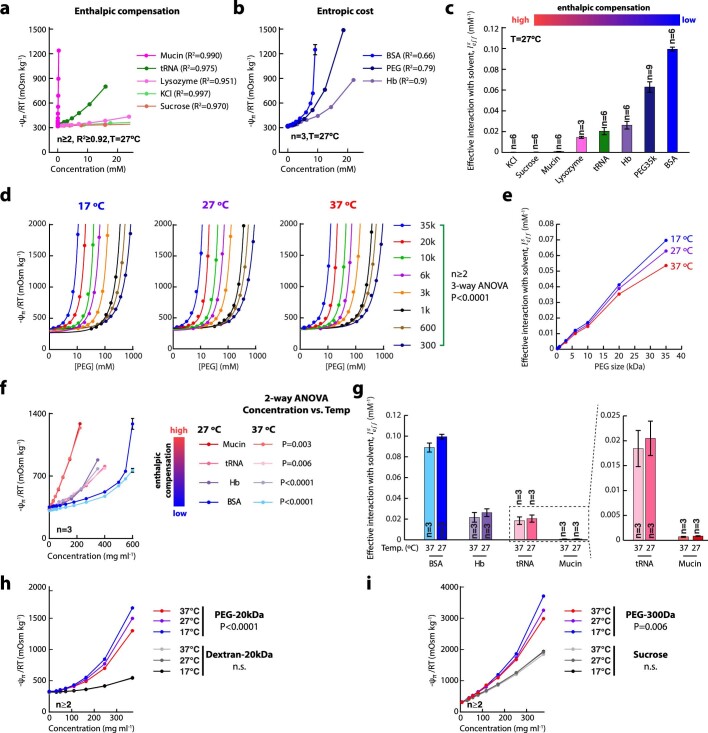
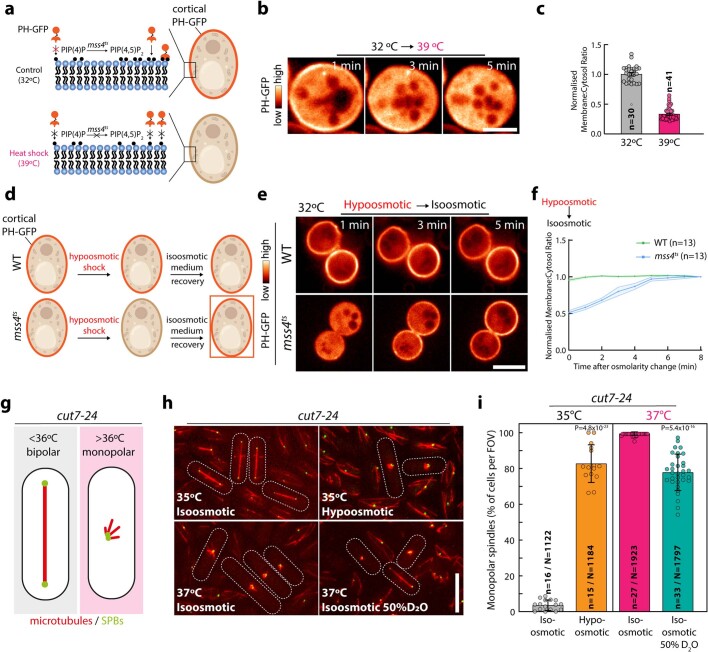
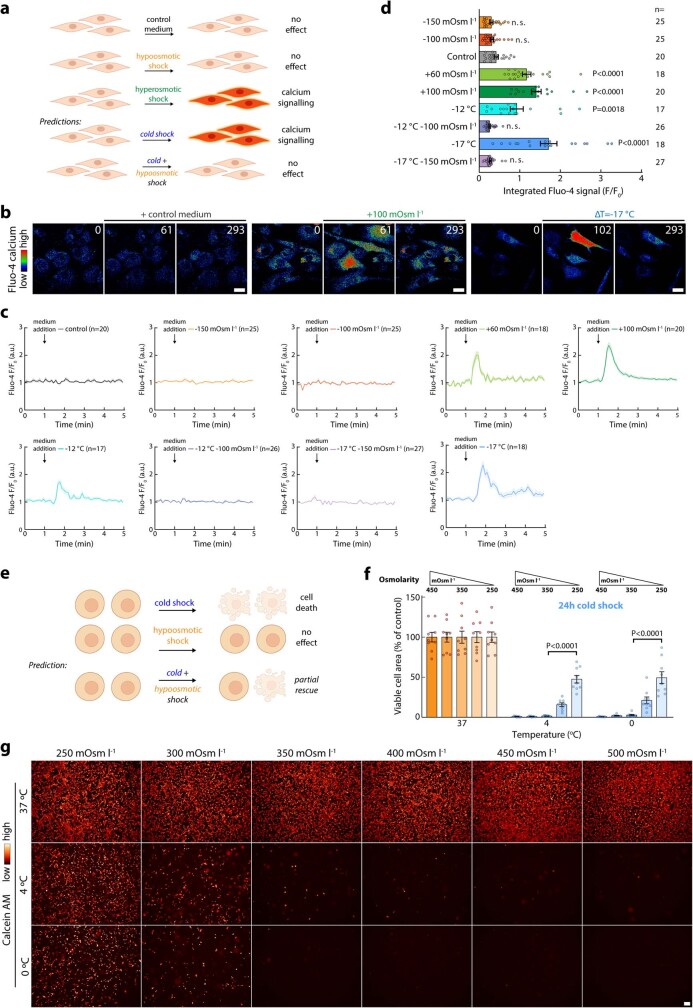
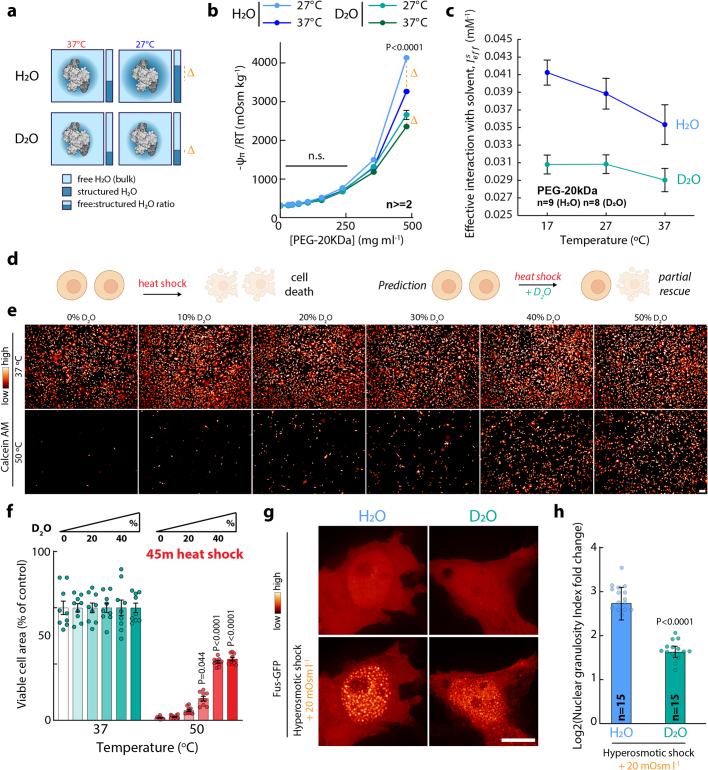
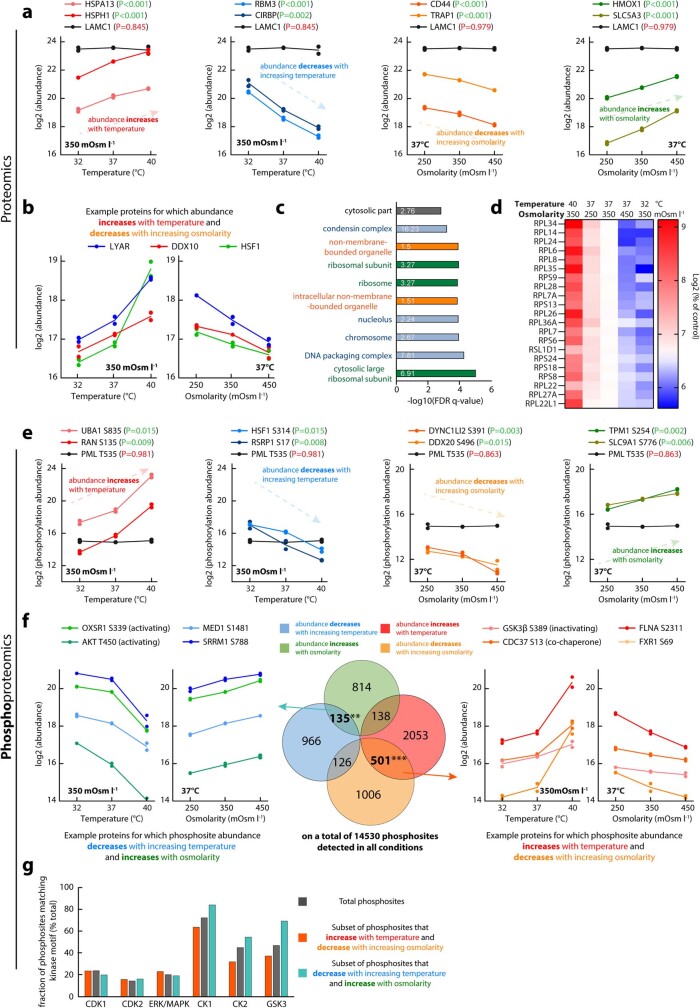
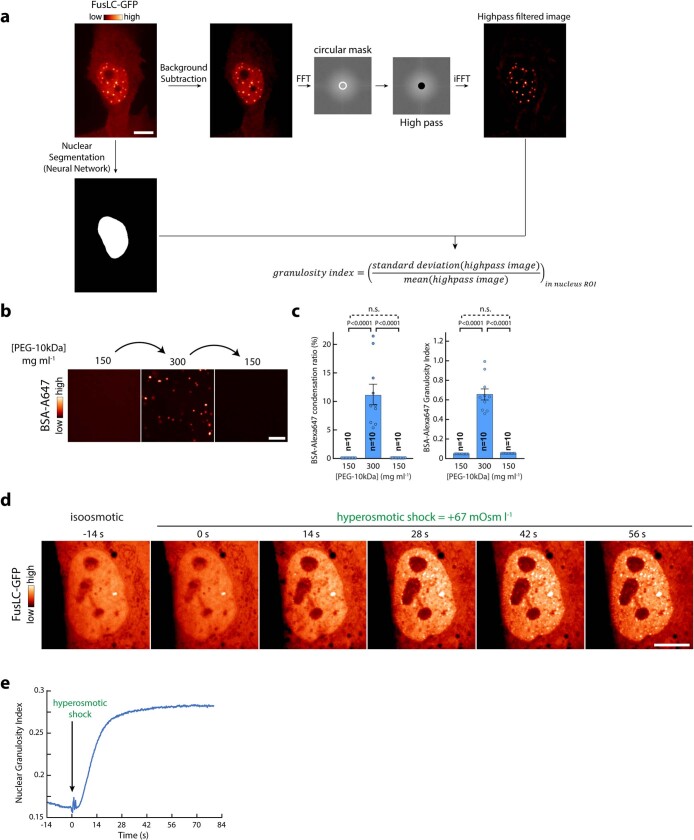
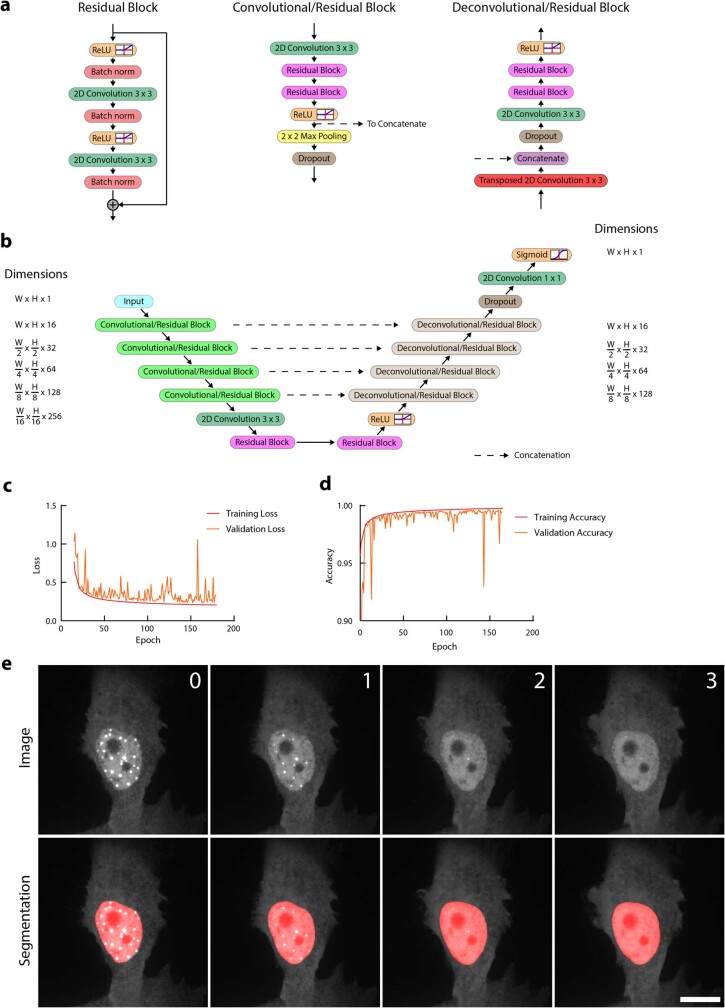
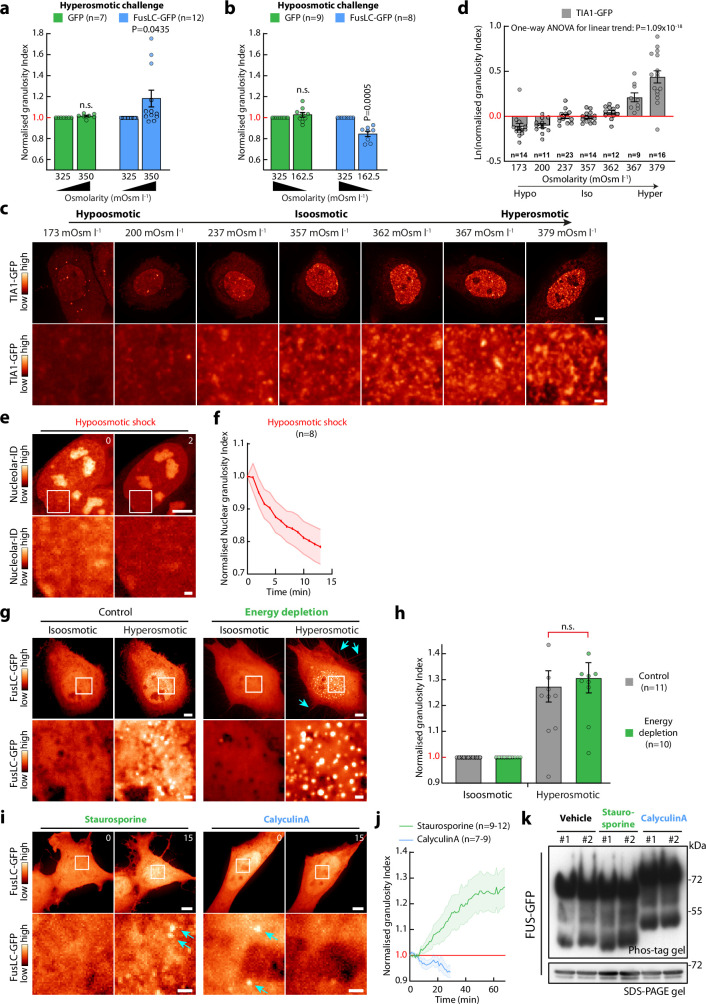
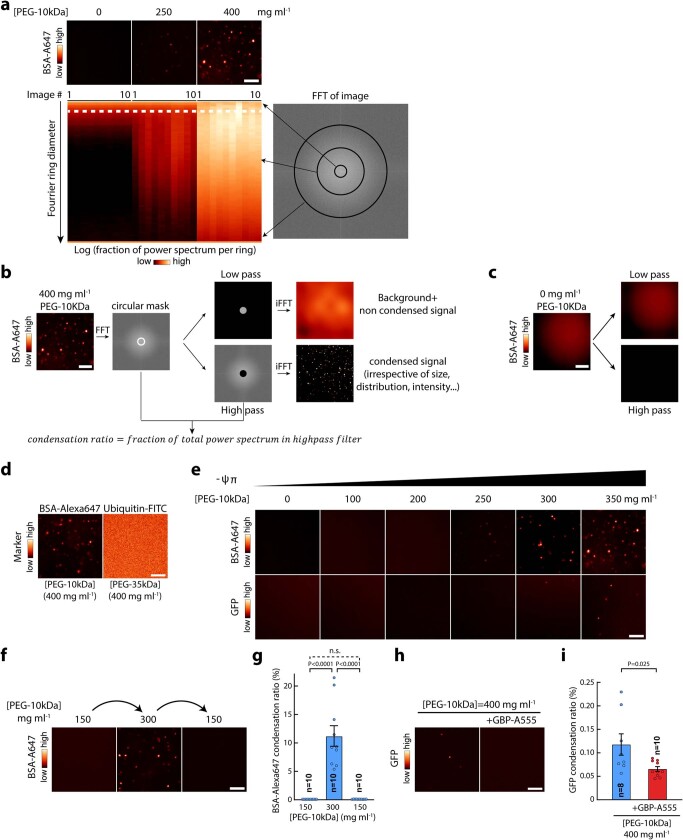
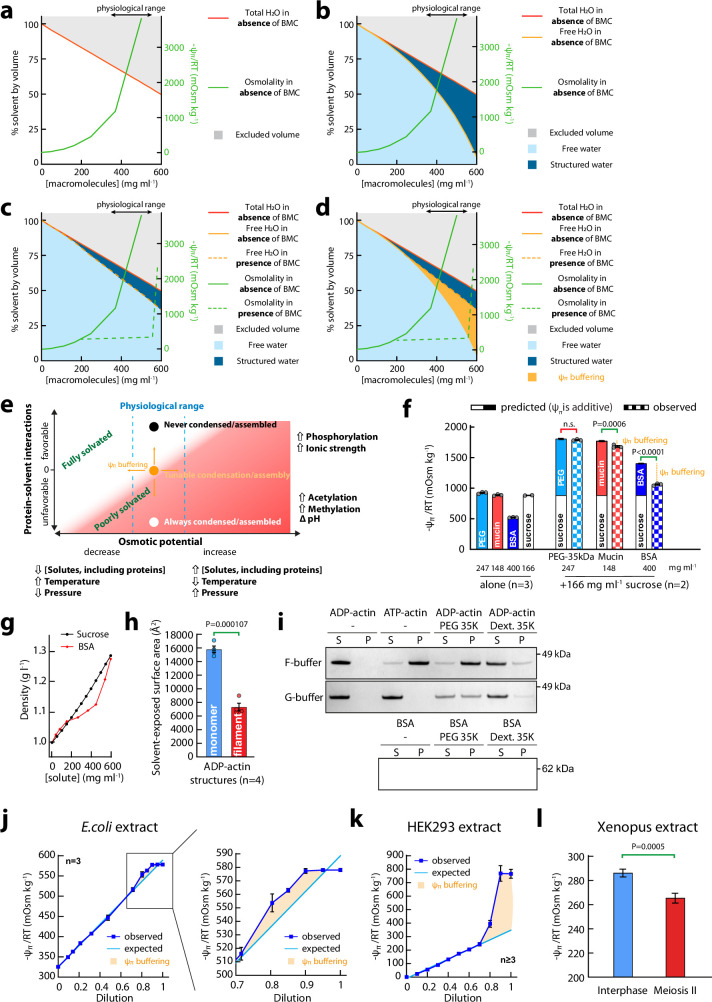
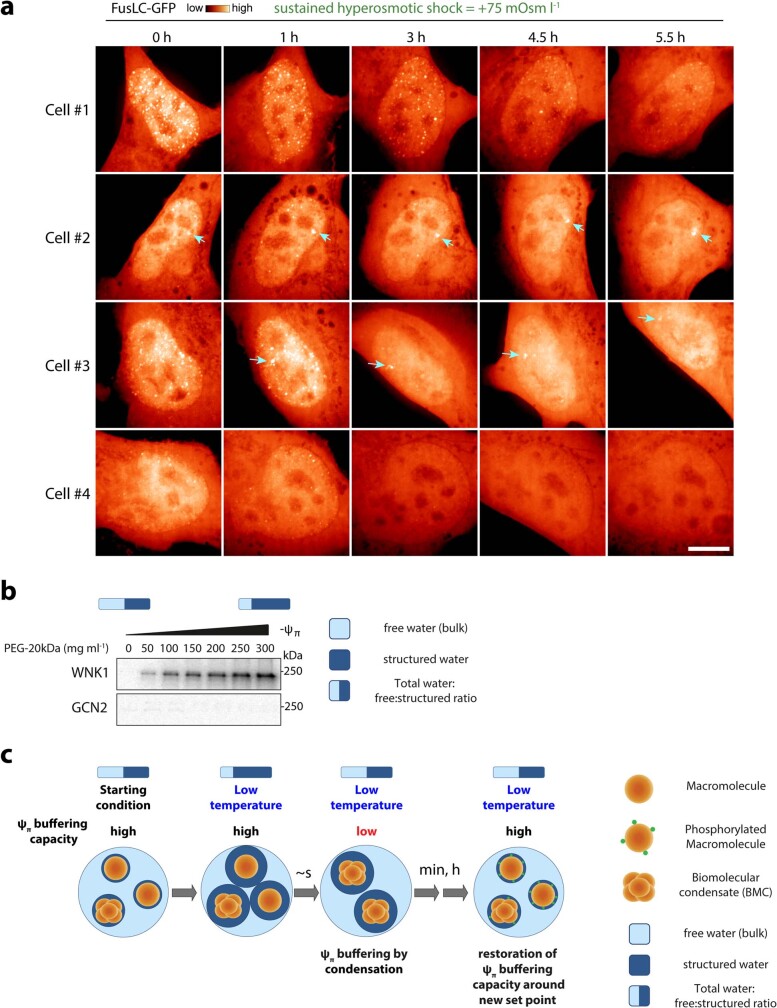
Optimum protein function and biochemical activity critically depends on water availability because solvent thermodynamics drive protein folding and macromolecular interactions1. Reciprocally, macromolecules restrict the movement of 'structured' water molecules within their hydration layers, reducing the available 'free' bulk solvent and therefore the total thermodynamic potential energy of water, or water potential. Here, within concentrated macromolecular solutions such as the cytosol, we found that modest changes in temperature greatly affect the water potential, and are counteracted by opposing changes in osmotic strength. This duality of temperature and osmotic strength enables simple manipulations of solvent thermodynamics to prevent cell death after extreme cold or heat shock. Physiologically, cells must sustain their activity against fluctuating temperature, pressure and osmotic strength, which impact water availability within seconds. Yet, established mechanisms of water homeostasis act over much slower timescales2,3; we therefore postulated the existence of a rapid compensatory response. We find that this function is performed by water potential-driven changes in macromolecular assembly, particularly biomolecular condensation of intrinsically disordered proteins. The formation and dissolution of biomolecular condensates liberates and captures free water, respectively, quickly counteracting thermal or osmotic perturbations of water potential, which is consequently robustly buffered in the cytoplasm. Our results indicate that biomolecular condensation constitutes an intrinsic biophysical feedback response that rapidly compensates for intracellular osmotic and thermal fluctuations. We suggest that preserving water availability within the concentrated cytosol is an overlooked evolutionary driver of protein (dis)order and function.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Protein condensation regulates water availability in cells.Nature. 2023 Nov;623(7988):698-699. doi: 10.1038/d41586-023-03098-z. Nature. 2023. PMID: 37853191 No abstract available.
References
-
- Rego, N. B. & Patel, A. J. Understanding hydrophobic effects: insights from water density fluctuations. Annu. Rev. Conden. Matter Phys.13, 303–324 (2022).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
