

Elucidating the cellular determinants of targeted membrane protein degradation by lysosome-targeting chimeras
- PMID: 37856615
- PMCID: PMC10766146
- DOI: 10.1126/science.adf6249
Elucidating the cellular determinants of targeted membrane protein degradation by lysosome-targeting chimeras
Abstract
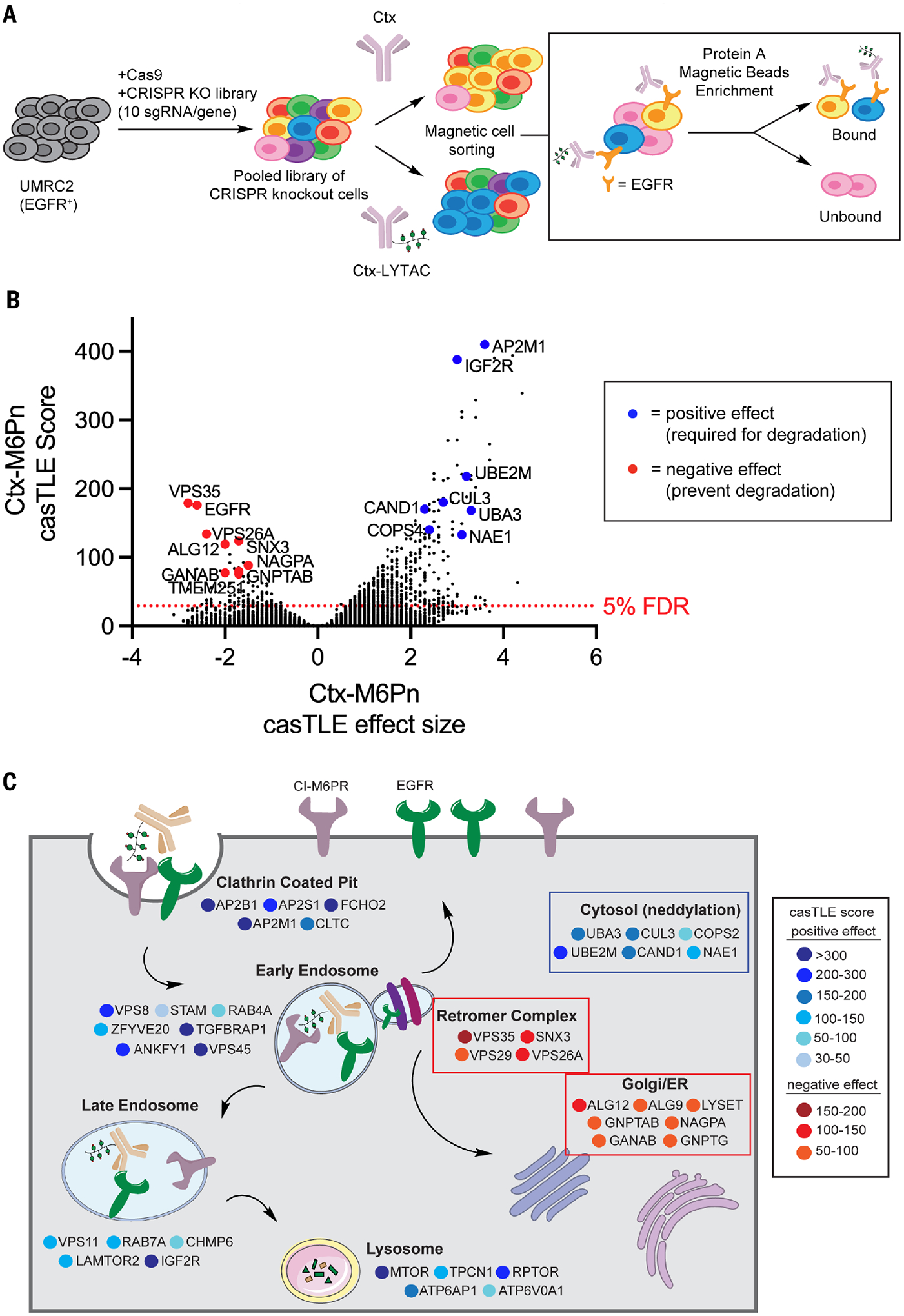
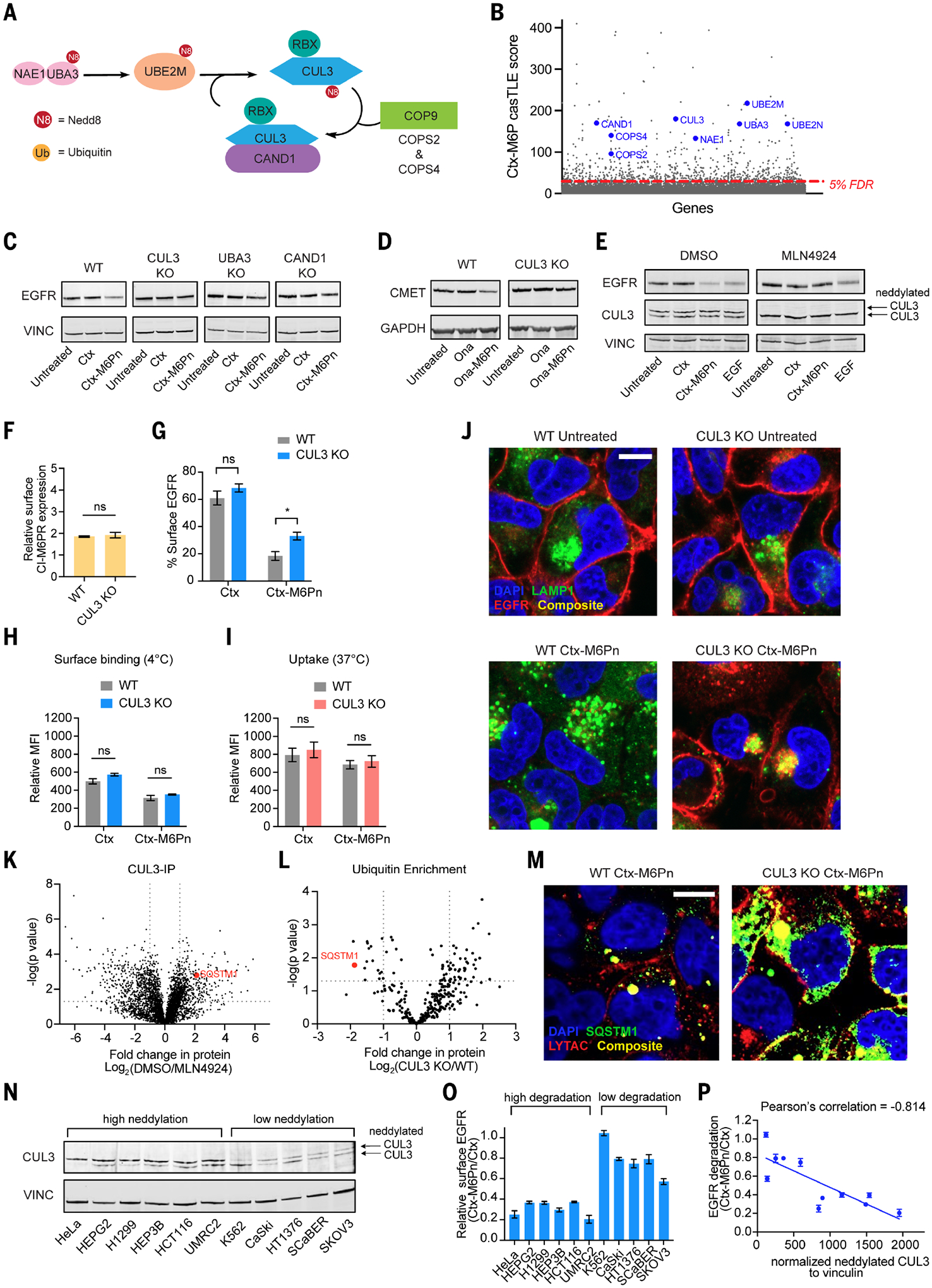
Targeted protein degradation can provide advantages over inhibition approaches in the development of therapeutic strategies. Lysosome-targeting chimeras (LYTACs) harness receptors, such as the cation-independent mannose 6-phosphate receptor (CI-M6PR), to direct extracellular proteins to lysosomes. In this work, we used a genome-wide CRISPR knockout approach to identify modulators of LYTAC-mediated membrane protein degradation in human cells. We found that disrupting retromer genes improved target degradation by reducing LYTAC recycling to the plasma membrane. Neddylated cullin-3 facilitated LYTAC-complex lysosomal maturation and was a predictive marker for LYTAC efficacy. A substantial fraction of cell surface CI-M6PR remains occupied by endogenous M6P-modified glycoproteins. Thus, inhibition of M6P biosynthesis increased the internalization of LYTAC-target complexes. Our findings inform design strategies for next-generation LYTACs and elucidate aspects of cell surface receptor occupancy and trafficking.
Conflict of interest statement
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
