

Drug repositioning strategy for the identification of novel telomere-damaging agents: A role for NAMPT inhibitors
- PMID: 37858982
- PMCID: PMC10652301
- DOI: 10.1111/acel.13944
Drug repositioning strategy for the identification of novel telomere-damaging agents: A role for NAMPT inhibitors
Abstract
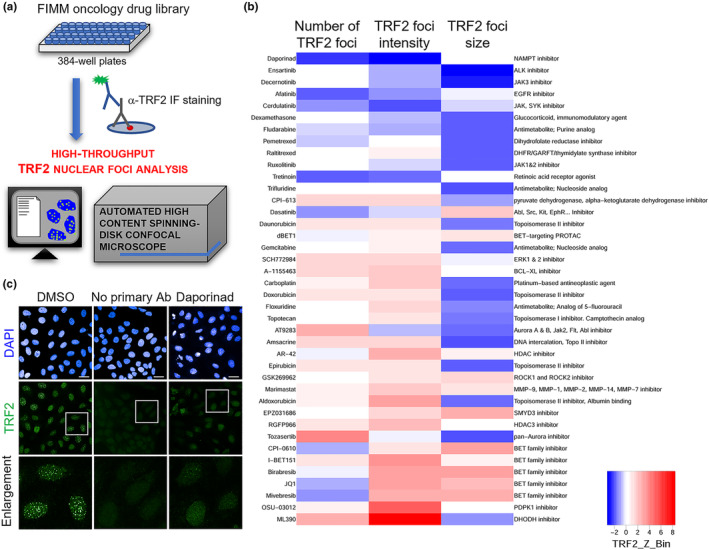
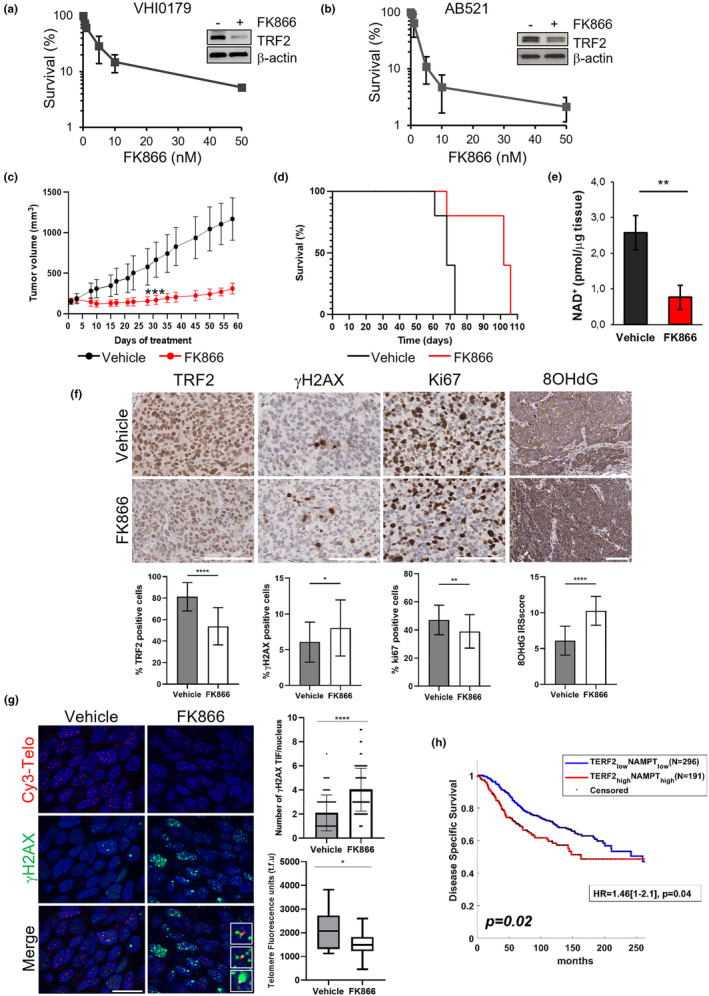
Drug repositioning strategy represents a valid tool to accelerate the pharmacological development through the identification of new applications for already existing compounds. In this view, we aimed at discovering molecules able to trigger telomere-localized DNA damage and tumor cell death. By applying an automated high-content spinning-disk microscopy, we performed a screening aimed at identifying, on a library of 527 drugs, molecules able to negatively affect the expression of TRF2, a key protein in telomere maintenance. FK866, resulting from the screening as the best candidate hit, was then validated at biochemical and molecular levels and the mechanism underlying its activity in telomere deprotection was elucidated both in vitro and in vivo. The results of this study allow us to discover a novel role of FK866 in promoting, through the production of reactive oxygen species, telomere loss and deprotection, two events leading to an accumulation of DNA damage and tumor cell death. The ability of FK866 to induce telomere damage and apoptosis was also demonstrated in advanced preclinical models evidencing the antitumoral activity of FK866 in triple-negative breast cancer-a particularly aggressive breast cancer subtype still orphan of targeted therapies and characterized by high expression levels of both NAMPT and TRF2. Overall, our findings pave the way to the development of novel anticancer strategies to counteract triple-negative breast cancer, based on the use of telomere deprotecting agents, including NAMPT inhibitors, that would rapidly progress from bench to bedside.
Keywords: NAMPT inhibitor; TRF2; anticancer therapy; cell death; drug screening; oxidative DNA damage; telomeres.
© 2023 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
None declared.
Figures




References
-
- Bejarano, L. , Bosso, G. , Louzame, J. , Serrano, R. , Gómez‐Casero, E. , Martínez‐Torrecuadrada, J. , Martínez, S. , Blanco‐Aparicio, C. , Pastor, J. , & Blasco, M. A. (2019). Multiple cancer pathways regulate telomere protection. EMBO Molecular Medicine, 11(7), e10292. 10.15252/EMMM.201910292 - DOI - PMC - PubMed
-
- Bejarano, L. , Schuhmacher, A. J. , Méndez, M. , Megías, D. , Blanco‐Aparicio, C. , Martínez, S. , Pastor, J. , Squatrito, M. , & Blasco, M. A. (2017). Inhibition of TRF1 telomere protein impairs tumor initiation and progression in glioblastoma mouse models and patient‐derived xenografts. Cancer Cell, 32(5), 590–607.e4. 10.1016/J.CCELL.2017.10.006 - DOI - PubMed
-
- Bruna, A. , Rueda, O. M. , Greenwood, W. , Batra, A. S. , Callari, M. , Batra, R. N. , Pogrebniak, K. , Sandoval, J. , Cassidy, J. W. , Tufegdzic‐Vidakovic, A. , Sammut, S. J. , Jones, L. , Provenzano, E. , Baird, R. , Eirew, P. , Hadfield, J. , Eldridge, M. , McLaren‐Douglas, A. , Barthorpe, A. , … Caldas, C. (2016). A biobank of breast cancer explants with preserved intra‐tumor heterogeneity to screen anticancer compounds. Cell, 167(1), 260–274.e22. 10.1016/j.cell.2016.08.041 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
