

The impact of molecular variants, crystallization conditions and the space group on ligand-protein complexes: a case study on bacterial phosphotriesterase
- PMID: 37860961
- PMCID: PMC10619419
- DOI: 10.1107/S2059798323007672
The impact of molecular variants, crystallization conditions and the space group on ligand-protein complexes: a case study on bacterial phosphotriesterase
Abstract
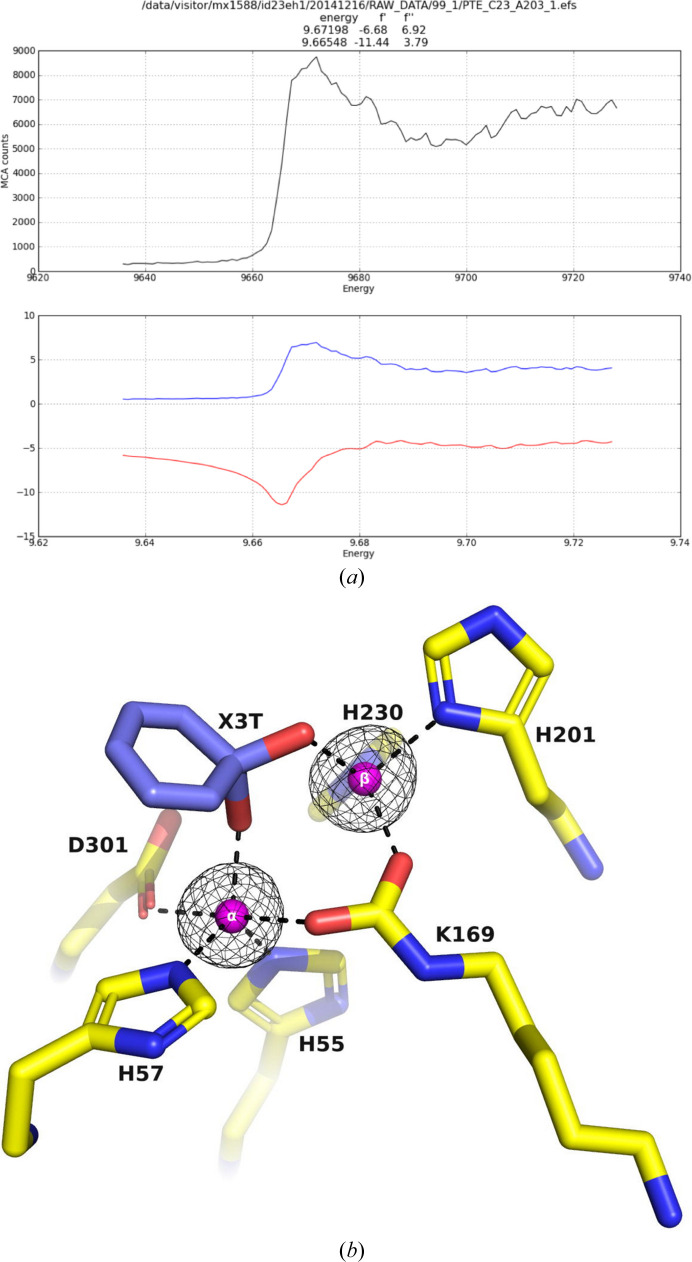
A bacterial phosphotriesterase was employed as an experimental paradigm to examine the effects of multiple factors, such as the molecular constructs, the ligands used during protein expression and purification, the crystallization conditions and the space group, on the visualization of molecular complexes of ligands with a target enzyme. In this case, the ligands used were organophosphates that are fragments of the nerve agents and insecticides on which the enzyme acts as a bioscavenger. 12 crystal structures of various phosphotriesterase constructs obtained by directed evolution were analyzed, with resolutions of up to 1.38 Å. Both apo forms and holo forms, complexed with the organophosphate ligands, were studied. Crystals obtained from three different crystallization conditions, crystallized in four space groups, with and without N-terminal tags, were utilized to investigate the impact of these factors on visualizing the organophosphate complexes of the enzyme. The study revealed that the tags used for protein expression can lodge in the active site and hinder ligand binding. Furthermore, the space group in which the protein crystallizes can significantly impact the visualization of bound ligands. It was also observed that the crystallization precipitants can compete with, and even preclude, ligand binding, leading to false positives or to the incorrect identification of lead drug candidates. One of the co-crystallization conditions enabled the definition of the spaces that accommodate the substituents attached to the P atom of several products of organophosphate substrates after detachment of the leaving group. The crystal structures of the complexes of phosphotriesterase with the organophosphate products reveal similar short interaction distances of the two partially charged O atoms of the P-O bonds with the exposed β-Zn2+ ion and the buried α-Zn2+ ion. This suggests that both Zn2+ ions have a role in stabilizing the transition state for substrate hydrolysis. Overall, this study provides valuable insights into the challenges and considerations involved in studying the crystal structures of ligand-protein complexes, highlighting the importance of careful experimental design and rigorous data analysis in ensuring the accuracy and reliability of the resulting phosphotriesterase-organophosphate structures.
Keywords: bioscavengers; crystallization conditions; expression systems; organophosphates; phosphotriesterases; space groups; zinc ions.
open access.
Figures











References
-
- Agirre, J., Atanasova, M., Bagdonas, H., Ballard, C. B., Baslé, A., Beilsten-Edmands, J., Borges, R. J., Brown, D. G., Burgos-Mármol, J. J., Berrisford, J. M., Bond, P. S., Caballero, I., Catapano, L., Chojnowski, G., Cook, A. G., Cowtan, K. D., Croll, T. I., Debreczeni, J. É., Devenish, N. E., Dodson, E. J., Drevon, T. R., Emsley, P., Evans, G., Evans, P. R., Fando, M., Foadi, J., Fuentes-Montero, L., Garman, E. F., Gerstel, M., Gildea, R. J., Hatti, K., Hekkelman, M. L., Heuser, P., Hoh, S. W., Hough, M. A., Jenkins, H. T., Jiménez, E., Joosten, R. P., Keegan, R. M., Keep, N., Krissinel, E. B., Kolenko, P., Kovalevskiy, O., Lamzin, V. S., Lawson, D. M., Lebedev, A. A., Leslie, A. G. W., Lohkamp, B., Long, F., Malý, M., McCoy, A. J., McNicholas, S. J., Medina, A., Millán, C., Murray, J. W., Murshudov, G. N., Nicholls, R. A., Noble, M. E. M., Oeffner, R., Pannu, N. S., Parkhurst, J. M., Pearce, N., Pereira, J., Perrakis, A., Powell, H. R., Read, R. J., Rigden, D. J., Rochira, W., Sammito, M., Sánchez Rodríguez, F., Sheldrick, G. M., Shelley, K. L., Simkovic, F., Simpkin, A. J., Skubak, P., Sobolev, E., Steiner, R. A., Stevenson, K., Tews, I., Thomas, J. M. H., Thorn, A., Valls, J. T., Uski, V., Usón, I., Vagin, A., Velankar, S., Vollmar, M., Walden, H., Waterman, D., Wilson, K. S., Winn, M. D., Winter, G., Wojdyr, M. & Yamashita, K. (2023). Acta Cryst. D79, 449–461.
-
- Aharoni, A., Amitai, G., Bernath, K., Magdassi, S. & Tawfik, D. S. (2005). Chem. Biol. 12, 1281–1289. - PubMed
-
- Benning, M. M., Hong, S. B., Raushel, F. M. & Holden, H. M. (2000). J. Biol. Chem. 275, 30556–30560. - PubMed
-
- Benning, M. M., Kuo, J. M., Raushel, F. M. & Holden, H. M. (1994). Biochemistry, 33, 15001–15007. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
