

Mitochondria-lysosome-related organelles mediate mitochondrial clearance during cellular dedifferentiation
- PMID: 37862166
- PMCID: PMC10842364
- DOI: 10.1016/j.celrep.2023.113291
Mitochondria-lysosome-related organelles mediate mitochondrial clearance during cellular dedifferentiation
Abstract
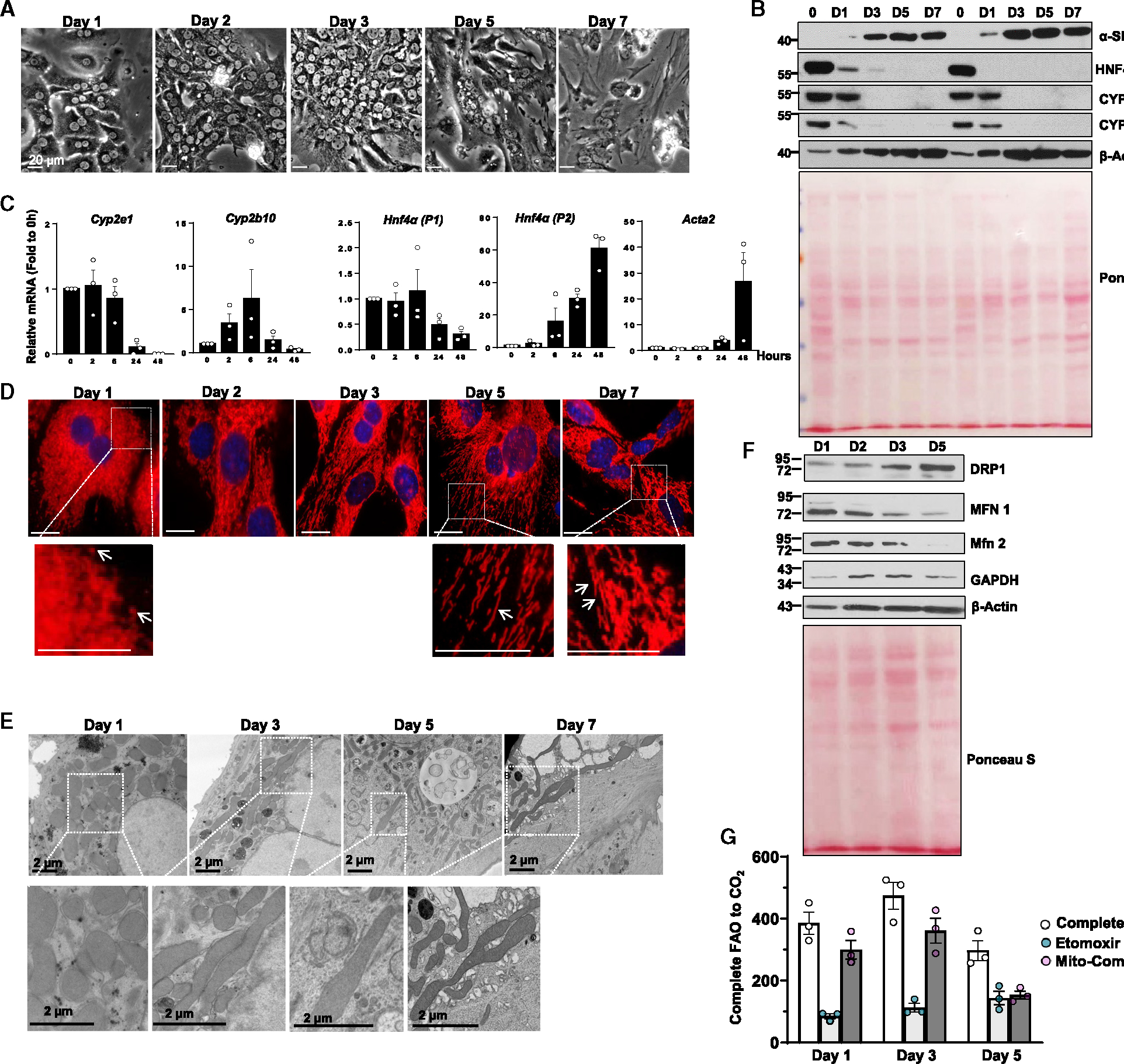
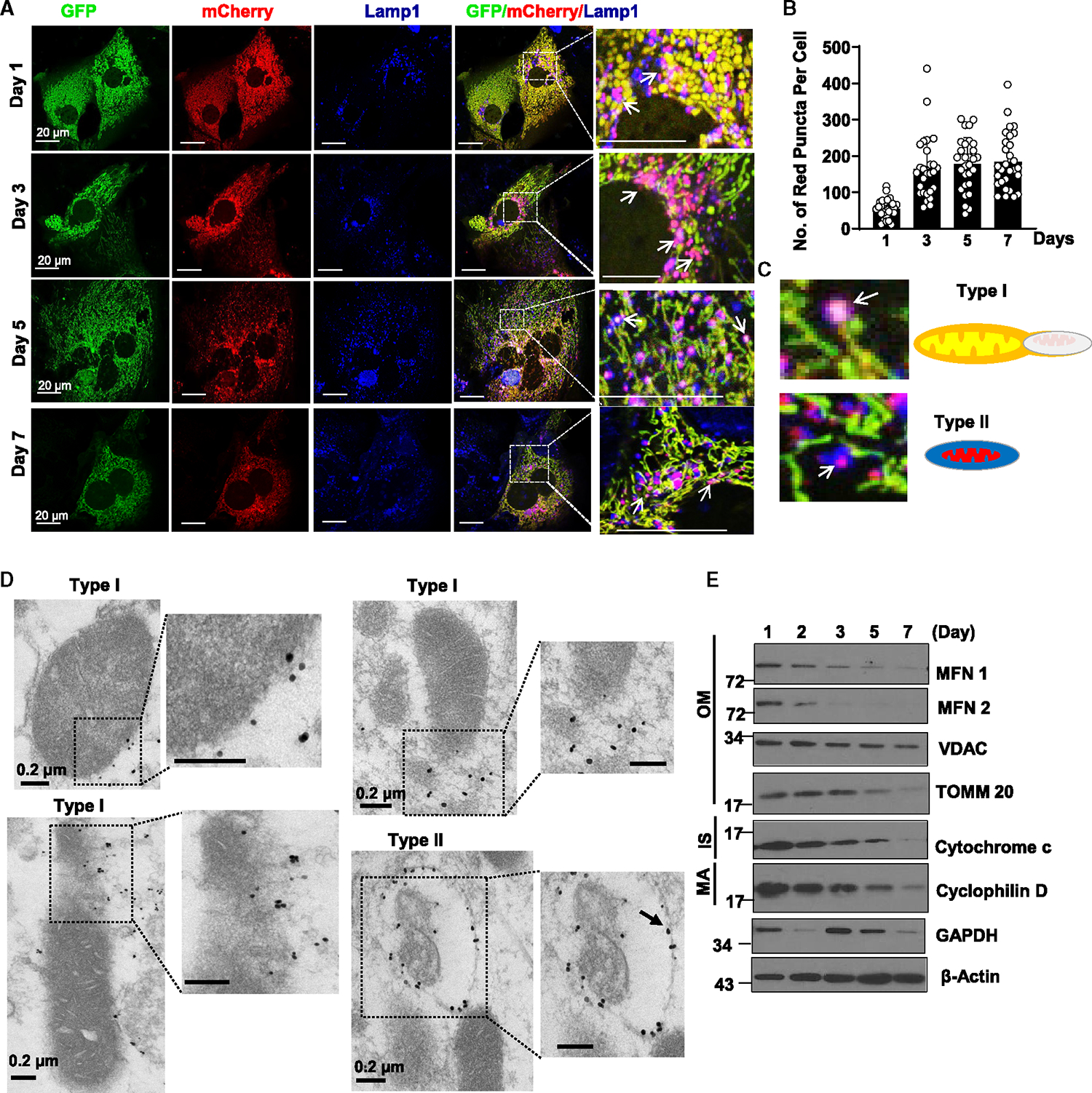
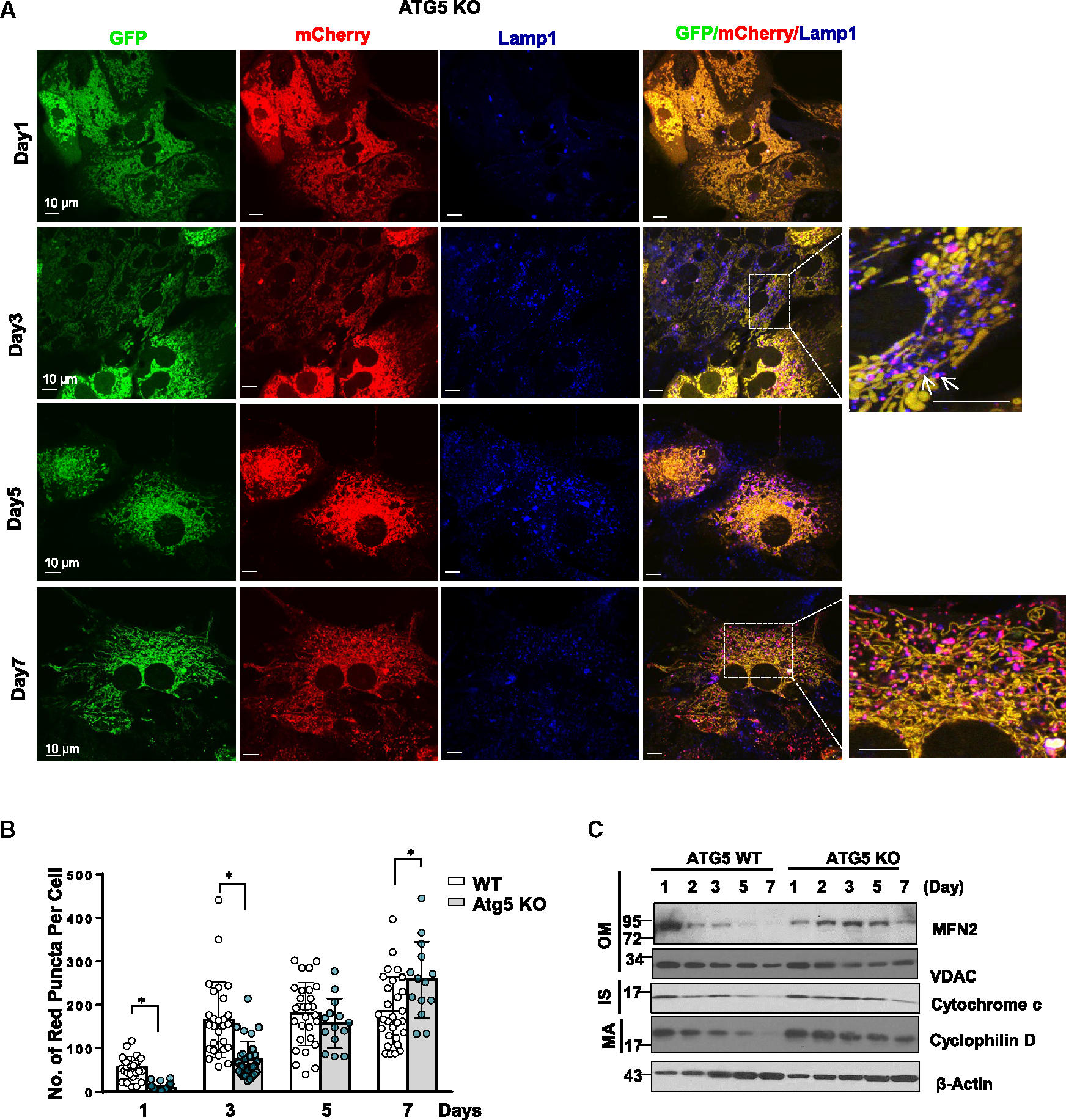
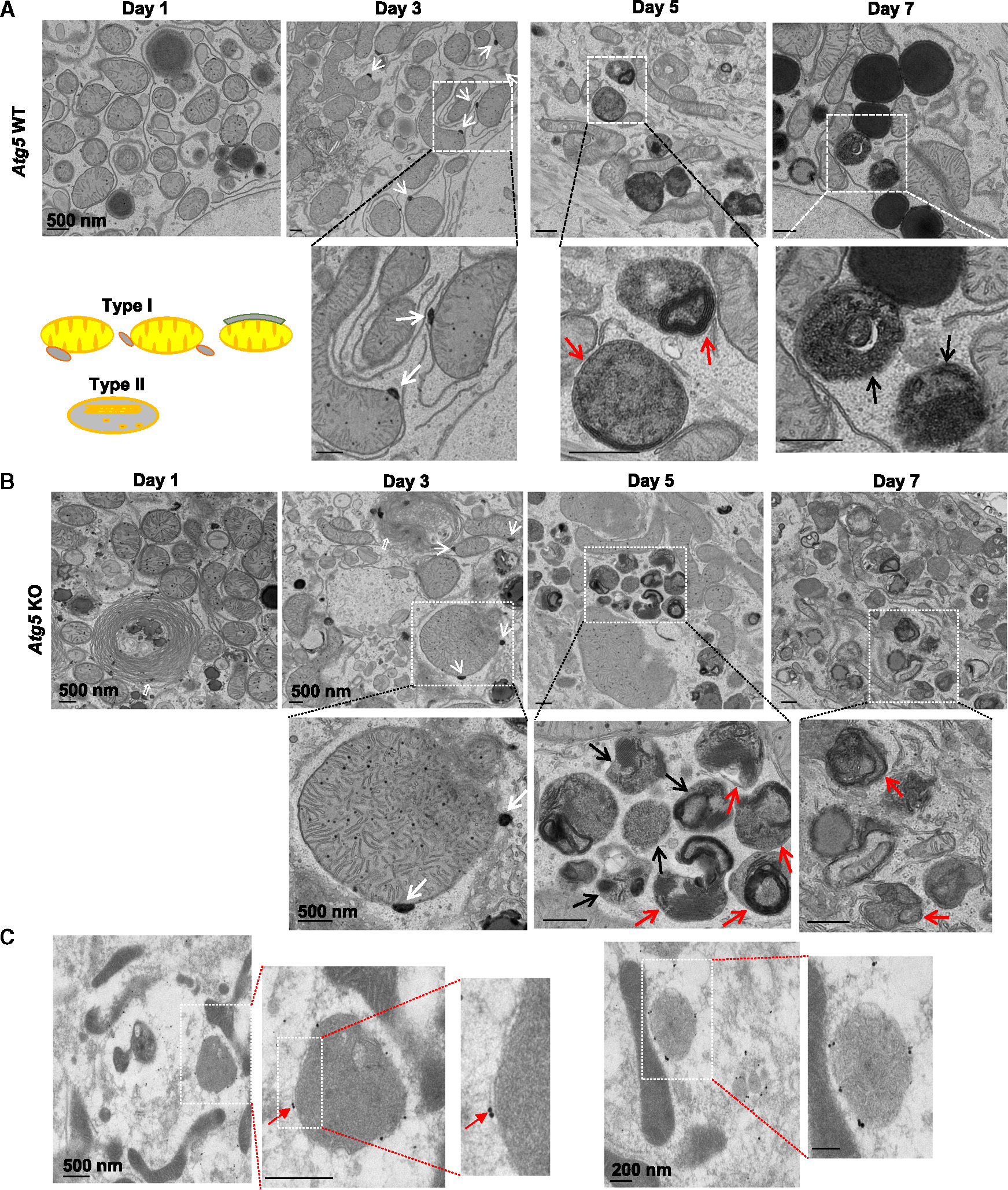
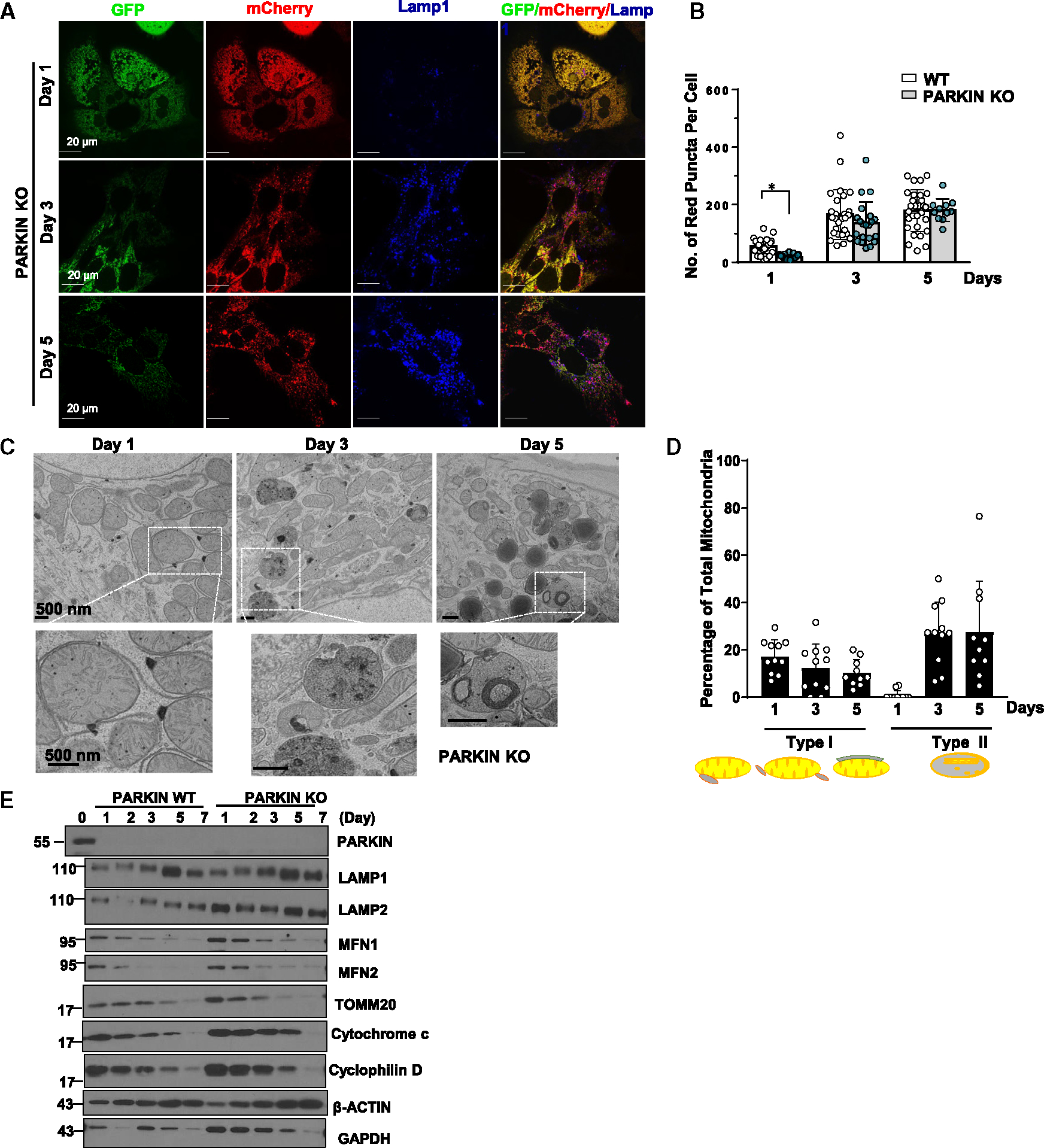
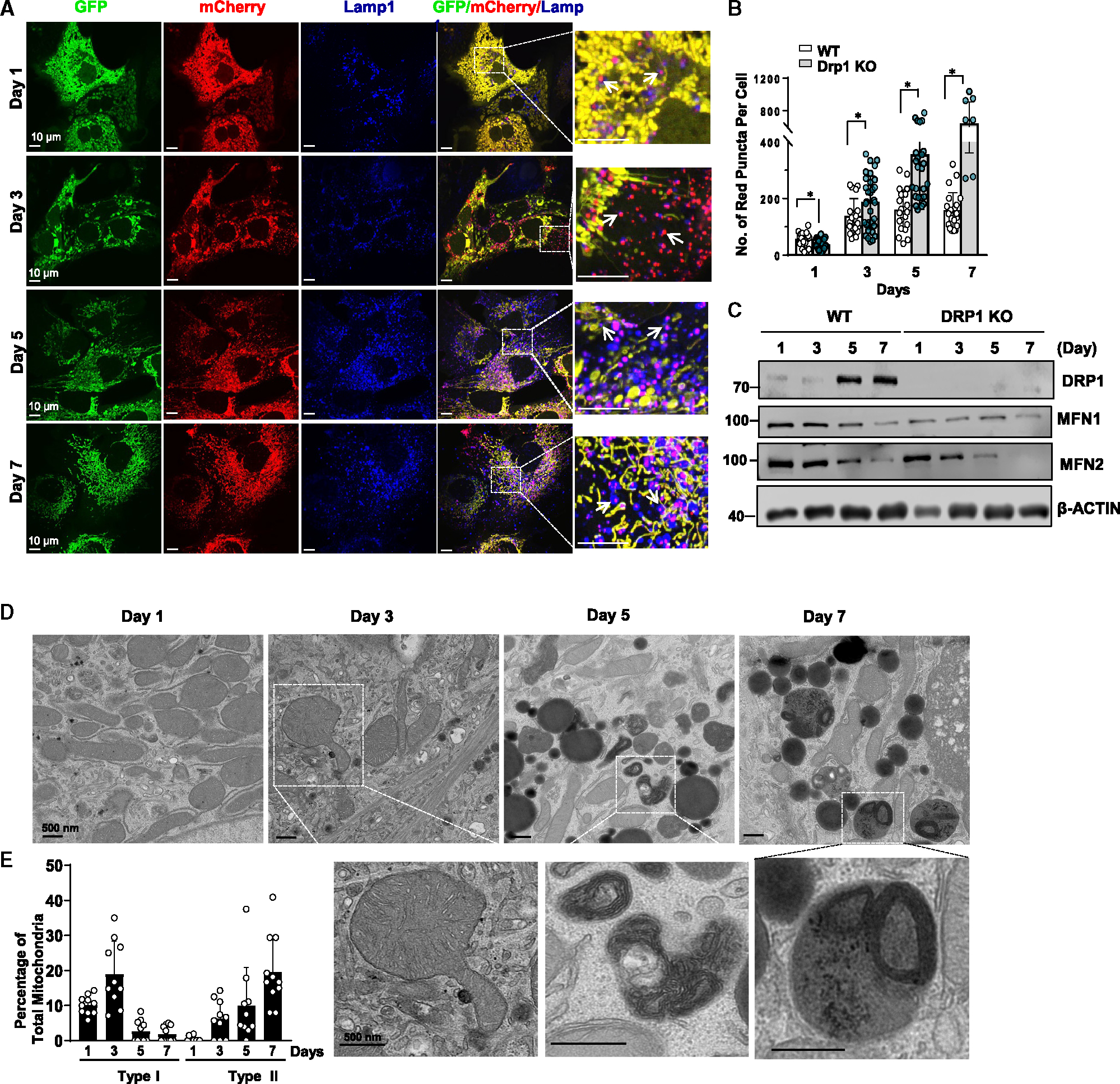
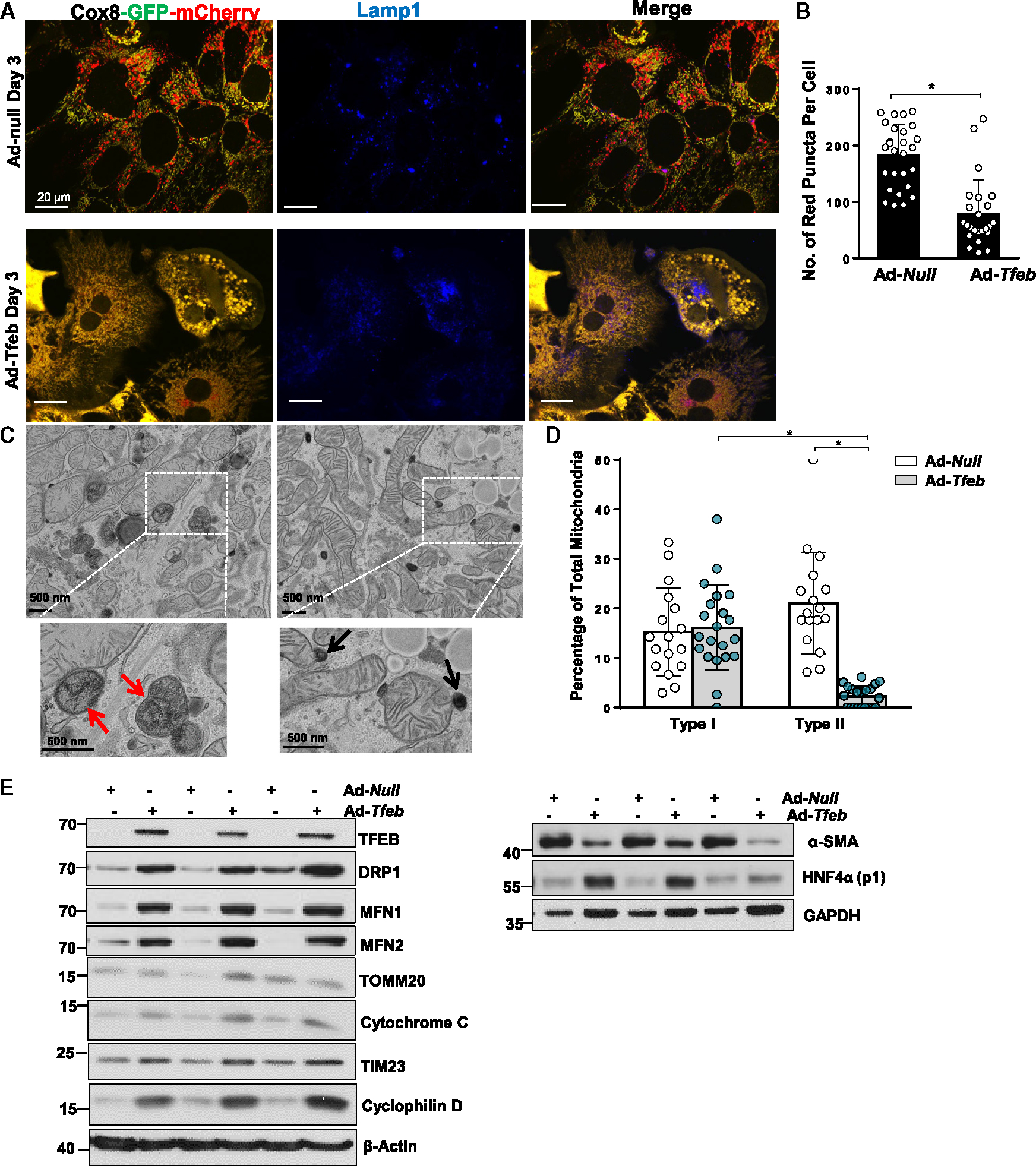
Dysfunctional mitochondria are removed via multiple pathways, such as mitophagy, a selective autophagy process. Here, we identify an intracellular hybrid mitochondria-lysosome organelle (termed the mitochondria-lysosome-related organelle [MLRO]), which regulates mitochondrial homeostasis independent of canonical mitophagy during hepatocyte dedifferentiation. The MLRO is an electron-dense organelle that has either a single or double membrane with both mitochondria and lysosome markers. Mechanistically, the MLRO is likely formed from the fusion of mitochondria-derived vesicles (MDVs) with lysosomes through a PARKIN-, ATG5-, and DRP1-independent process, which is negatively regulated by transcription factor EB (TFEB) and associated with mitochondrial protein degradation and hepatocyte dedifferentiation. The MLRO, which is galectin-3 positive, is reminiscent of damaged lysosome and could be cleared by overexpression of TFEB, resulting in attenuation of hepatocyte dedifferentiation. Together, results from this study suggest that the MLRO may act as an alternative mechanism for mitochondrial quality control independent of canonical autophagy/mitophagy involved in cell dedifferentiation.
Keywords: ATG5; CP: Cell biology; DRP1; autophagy; hepatocytes; lysosome; mitophagy.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
