

Cyclase-associated protein interacts with actin filament barbed ends to promote depolymerization and formin displacement
- PMID: 37863260
- PMCID: PMC10692737
- DOI: 10.1016/j.jbc.2023.105367
Cyclase-associated protein interacts with actin filament barbed ends to promote depolymerization and formin displacement
Abstract
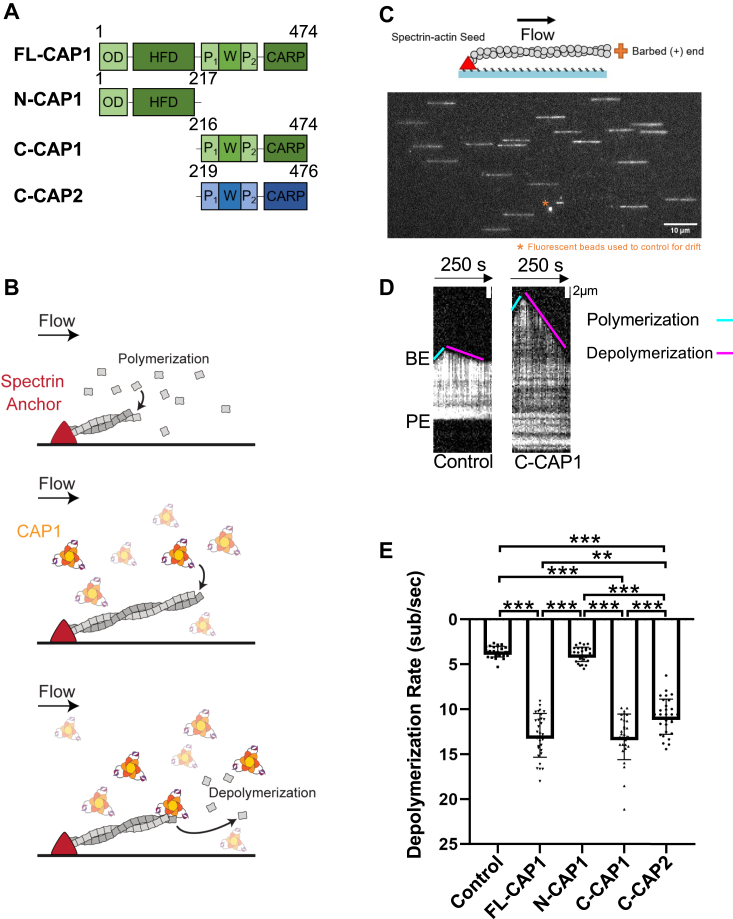
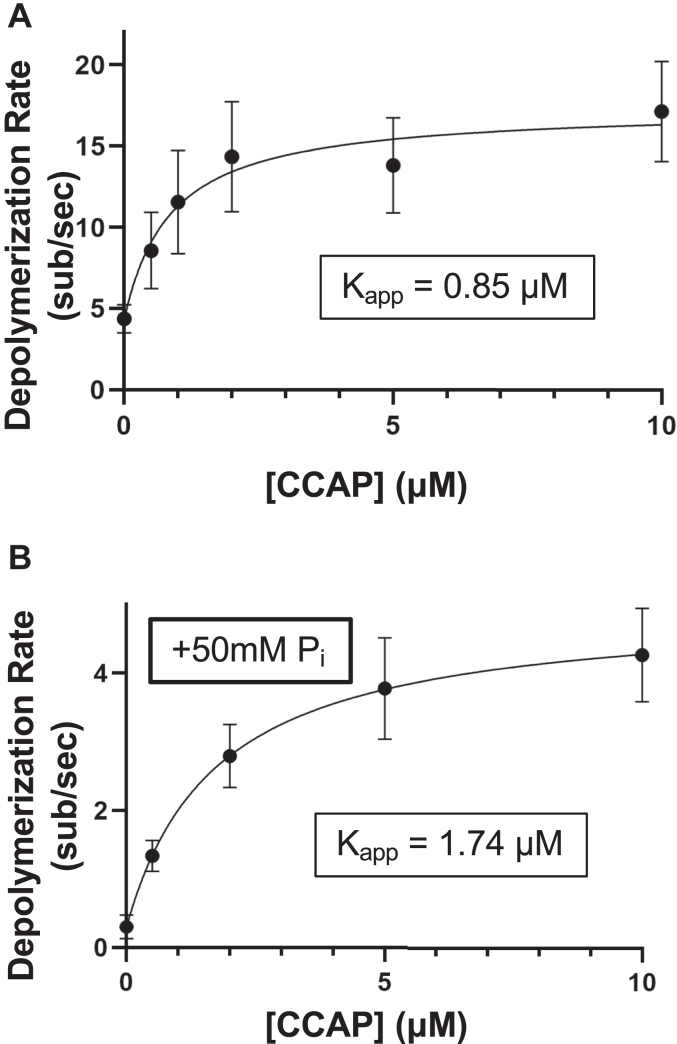
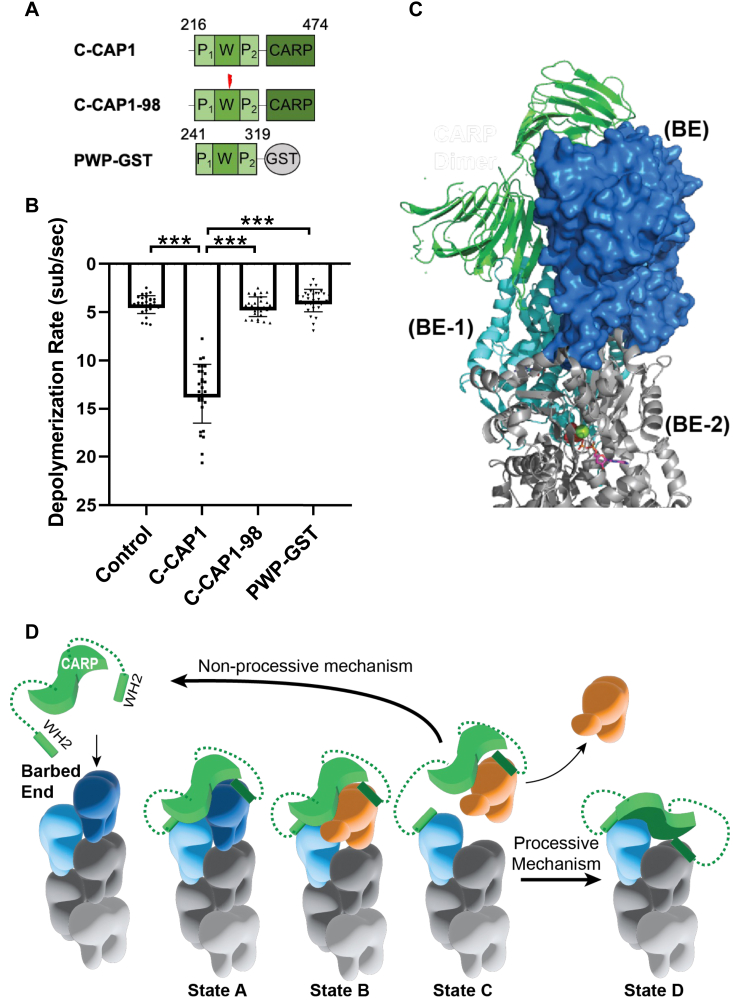
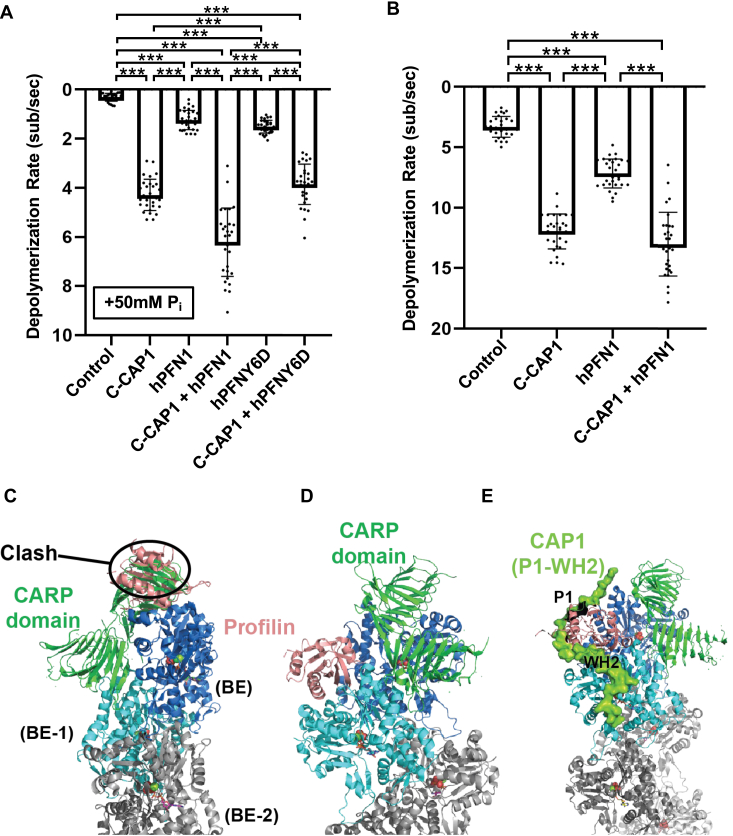
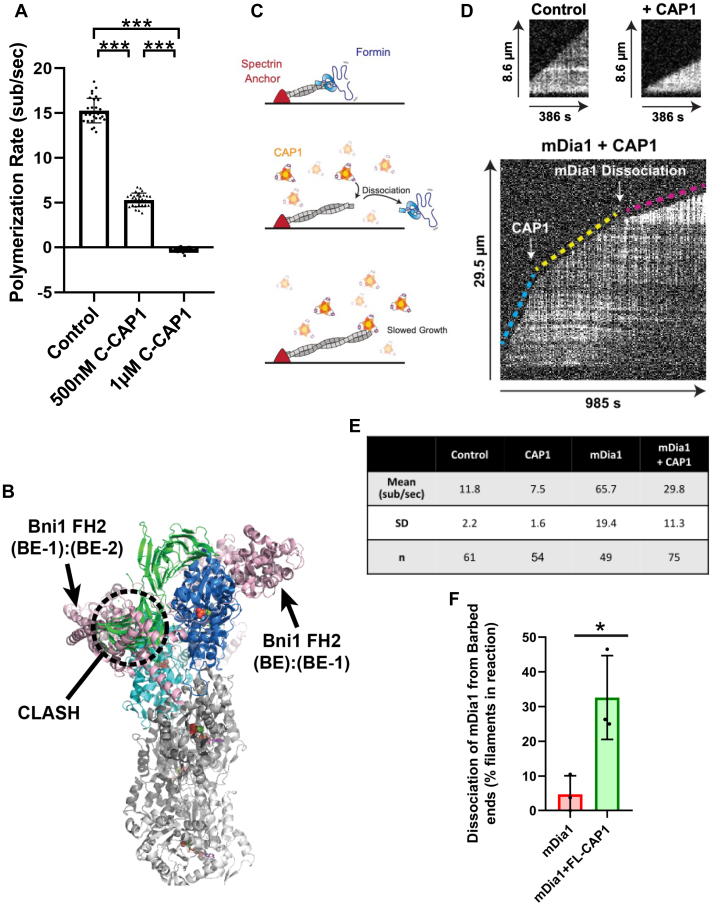
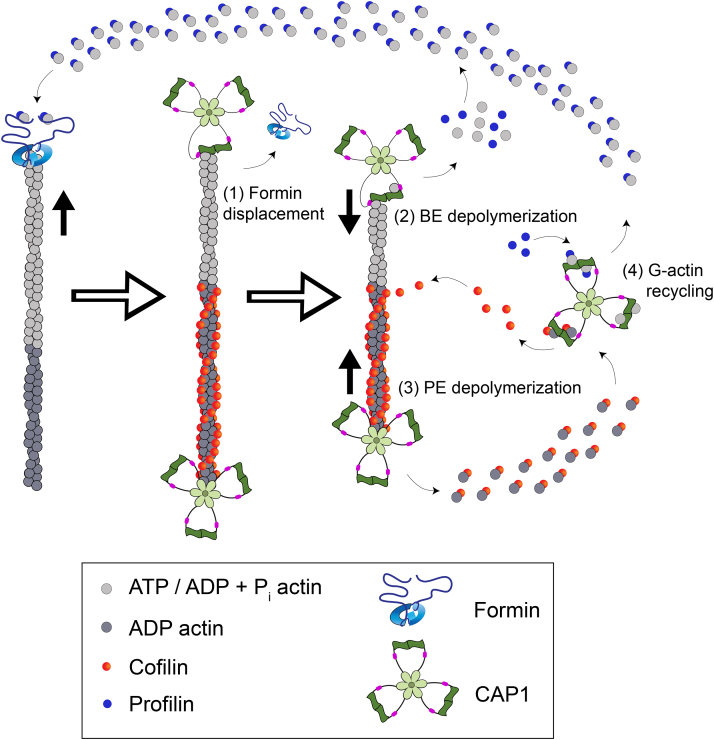
Cyclase-associated protein (CAP) has emerged as a central player in cellular actin turnover, but its molecular mechanisms of action are not yet fully understood. Recent studies revealed that the N terminus of CAP interacts with the pointed ends of actin filaments to accelerate depolymerization in conjunction with cofilin. Here, we use in vitro microfluidics-assisted TIRF microscopy to show that the C terminus of CAP promotes depolymerization at the opposite (barbed) ends of actin filaments. In the absence of actin monomers, full-length mouse CAP1 and C-terminal halves of CAP1 (C-CAP1) and CAP2 (C-CAP2) accelerate barbed end depolymerization. Using mutagenesis and structural modeling, we show that these activities are mediated by the WH2 and CARP domains of CAP. In addition, we observe that CAP collaborates with profilin to accelerate barbed end depolymerization and that these effects depend on their direct interaction, providing the first known example of CAP-profilin collaborative effects in regulating actin. In the presence of actin monomers, CAP1 attenuates barbed end growth and promotes formin dissociation. Overall, these findings demonstrate that CAP uses distinct domains and mechanisms to interact with opposite ends of actin filaments and drive turnover. Further, they contribute to the emerging view of actin barbed ends as sites of dynamic molecular regulation, where numerous proteins compete and cooperate with each other to tune polymer dynamics, similar to the rich complexity seen at microtubule ends.
Keywords: ADF; TIRF microscopy; actin; cofilin; cyclase-associated protein; turnover.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Pollard T.D., Borisy G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell. 2003;112:453–465. - PubMed
-
- Blanchoin L., Boujemaa-Paterski R., Sykes C., Plastino J. Actin dynamics, architecture, and mechanics in cell motility. Physiol. Rev. 2014;94:235–263. - PubMed
-
- Carlier M.-F., Shekhar S. Global treadmilling coordinates actin turnover and controls the size of actin networks. Nat. Rev. Mol. Cell Biol. 2017;18:389–401. - PubMed
-
- Chhabra E.S., Higgs H.N. The many faces of actin: matching assembly factors with cellular structures. Nat. Cell Biol. 2007;9:1110–1121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
