

TGFB1 induces fetal reprogramming and enhances intestinal regeneration
- PMID: 37865088
- PMCID: PMC10841757
- DOI: 10.1016/j.stem.2023.09.015
TGFB1 induces fetal reprogramming and enhances intestinal regeneration
Abstract
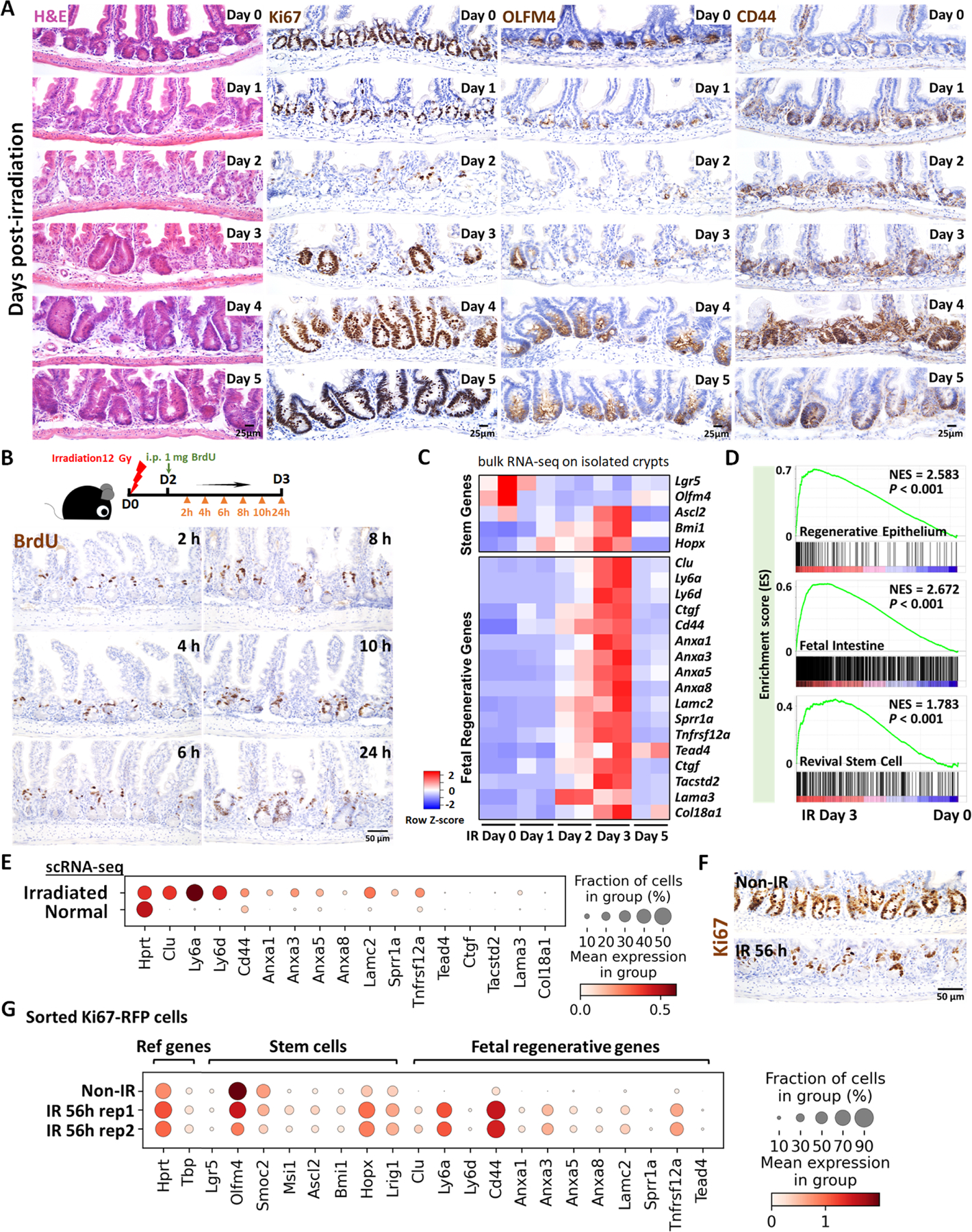
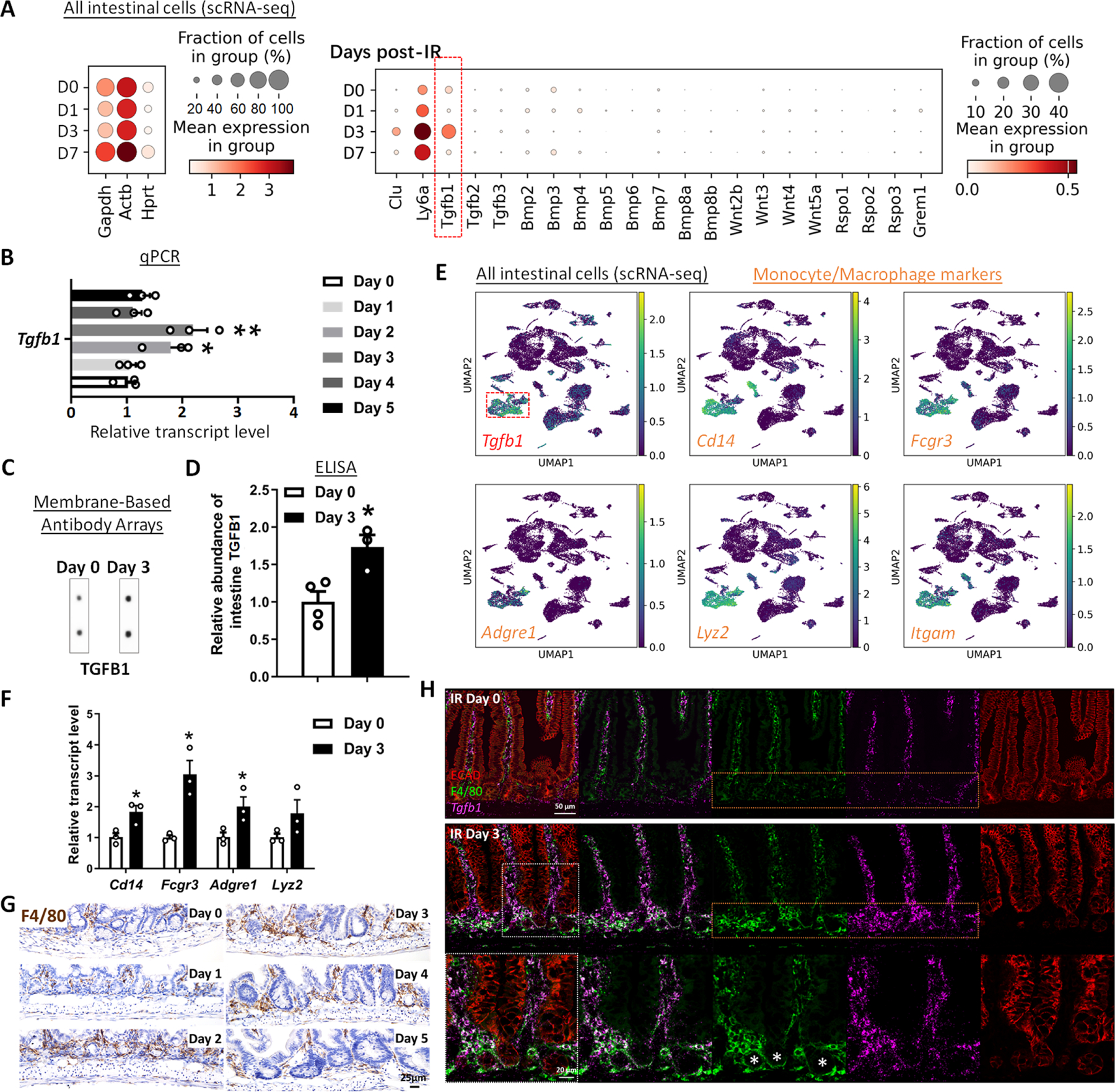
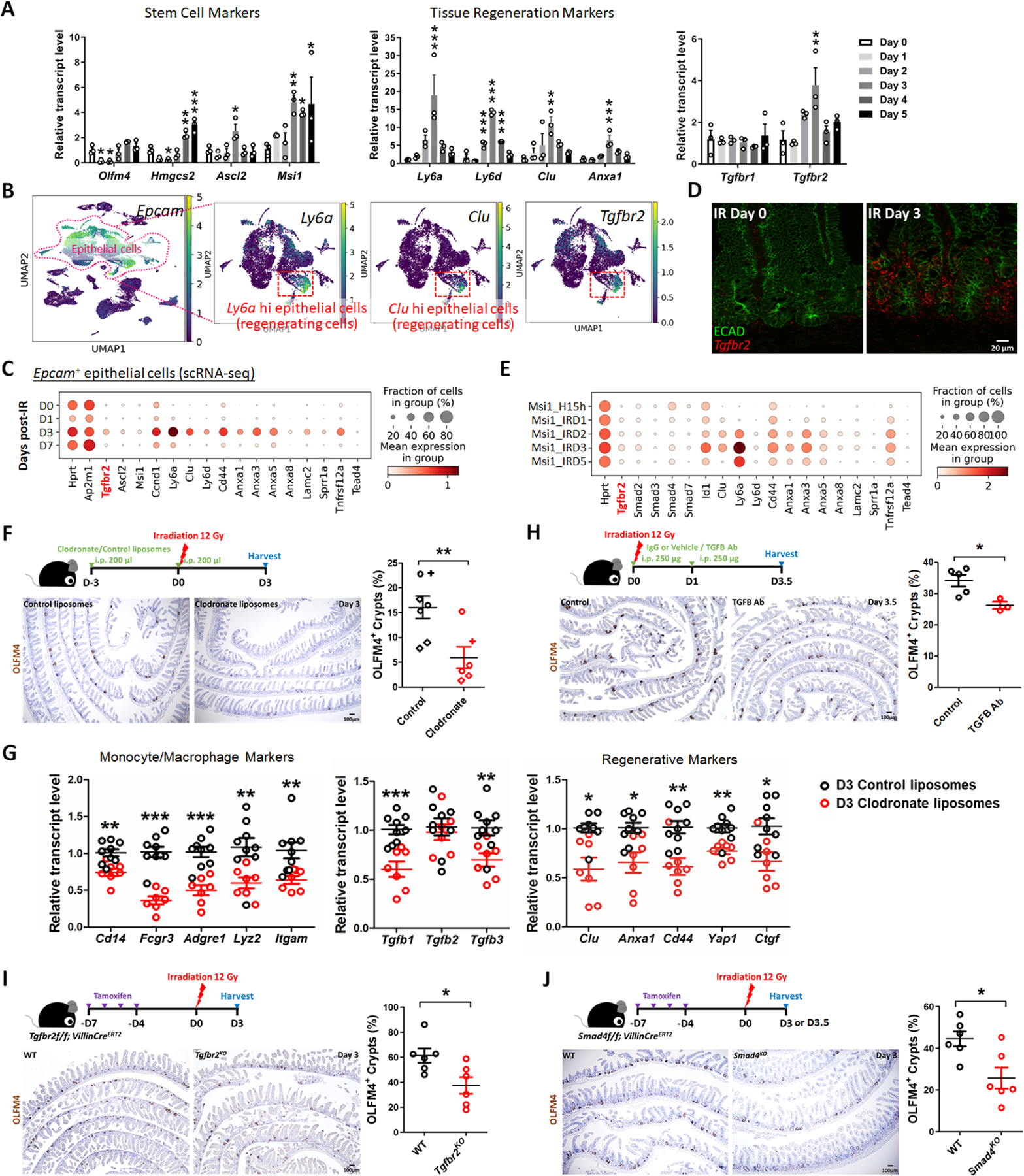
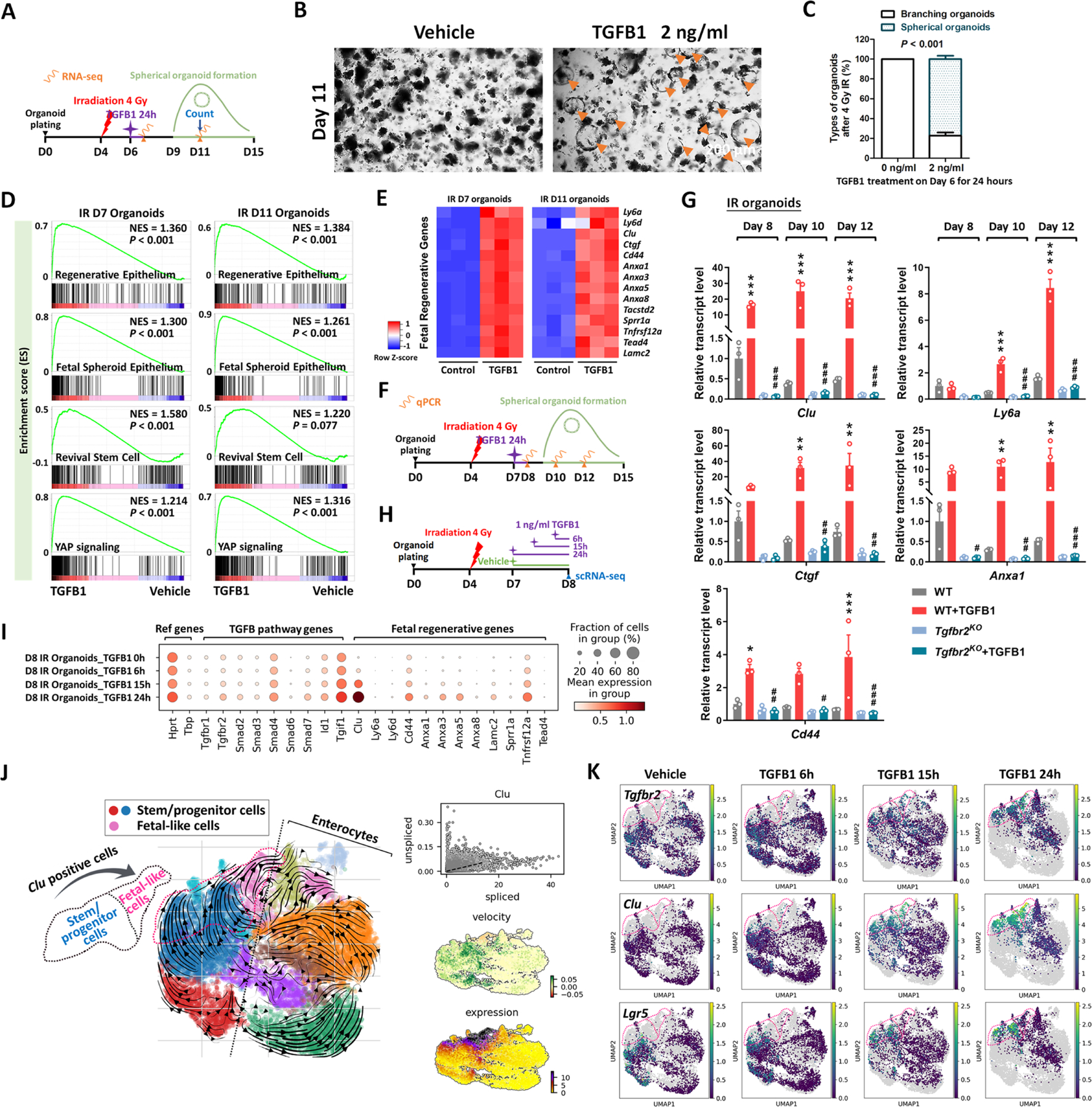
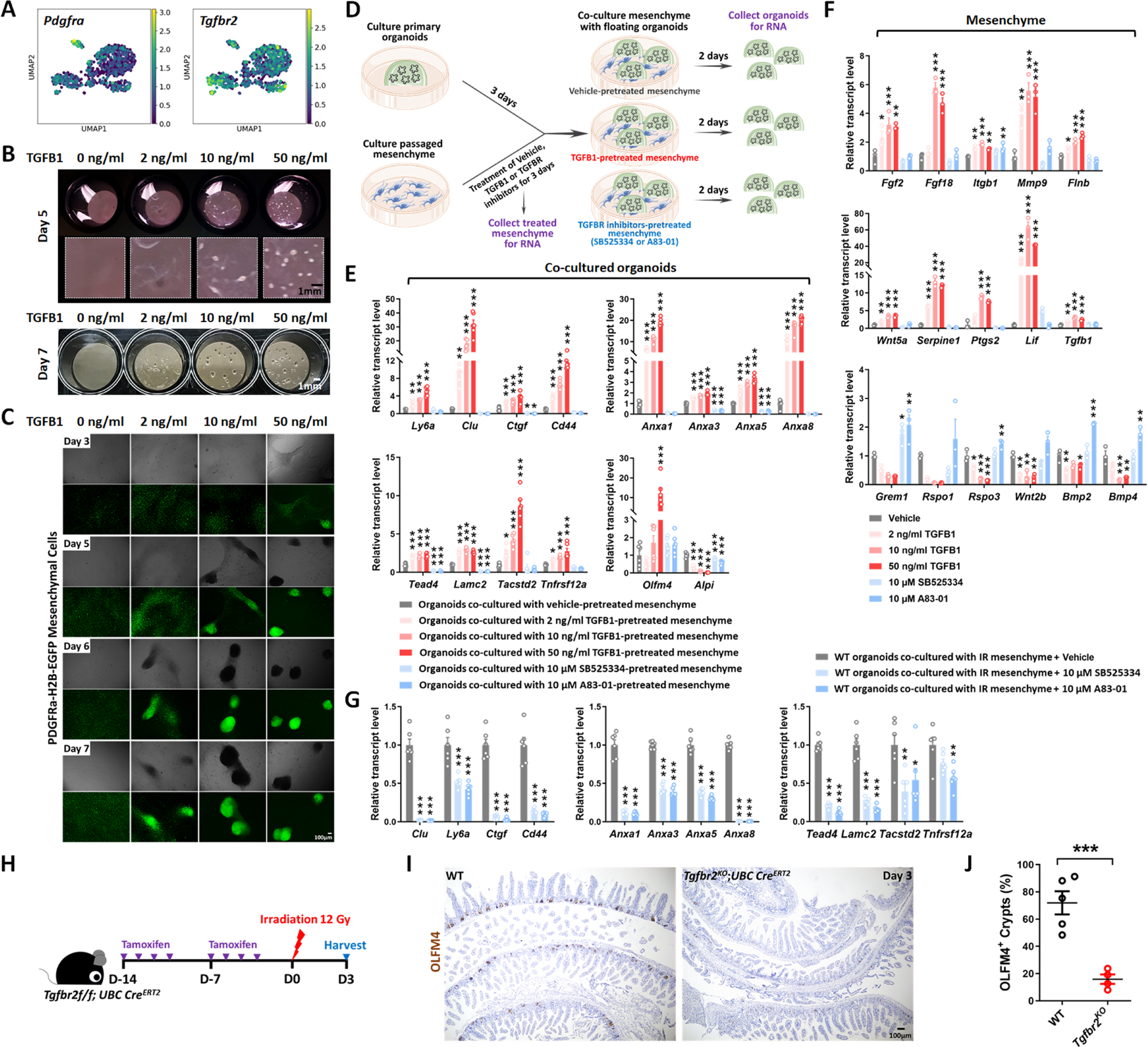
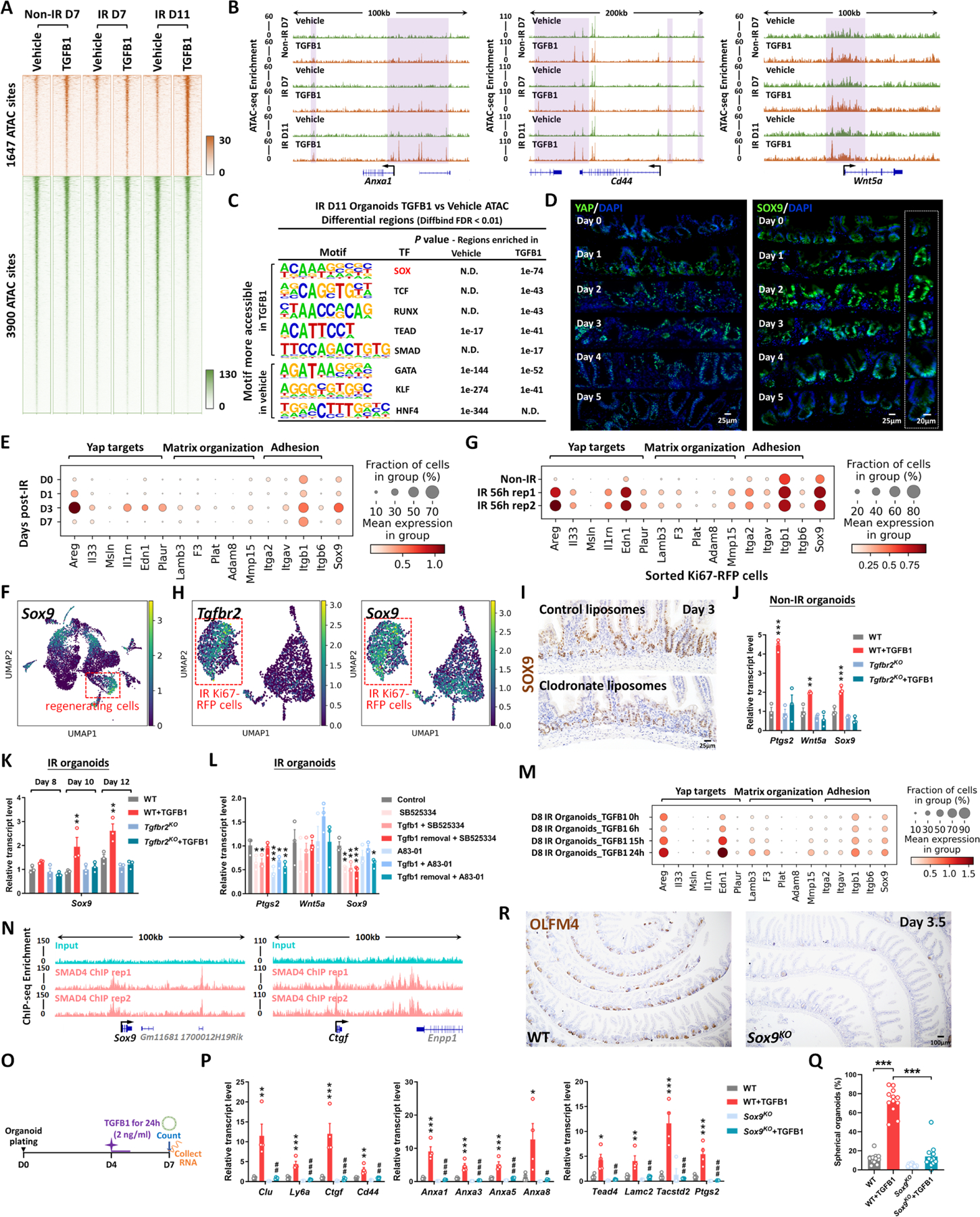
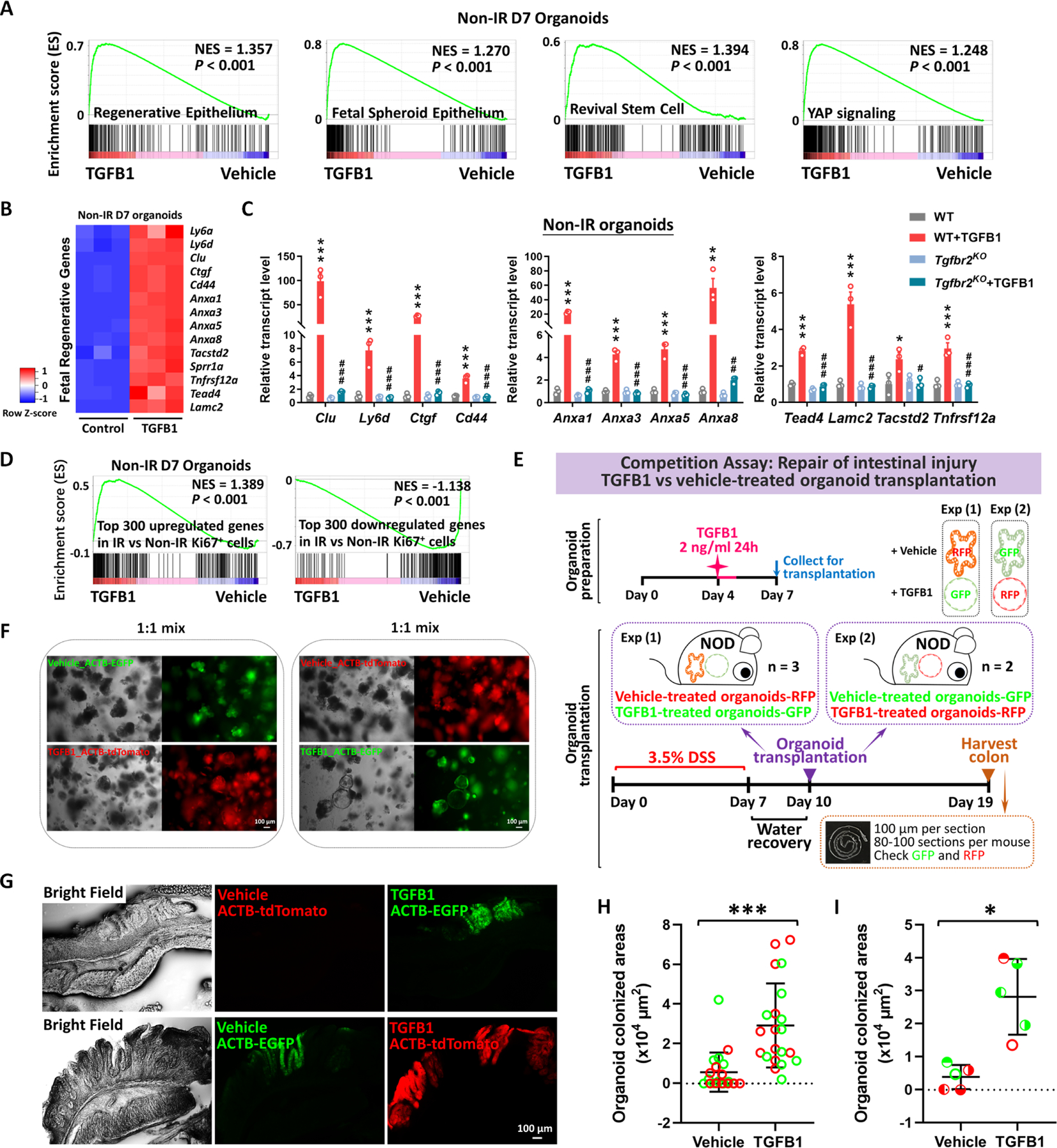
The gut epithelium has a remarkable ability to recover from damage. We employed a combination of high-throughput sequencing approaches, mouse genetics, and murine and human organoids and identified a role for TGFB signaling during intestinal regeneration following injury. At 2 days following irradiation (IR)-induced damage of intestinal crypts, a surge in TGFB1 expression is mediated by monocyte/macrophage cells at the location of damage. The depletion of macrophages or genetic disruption of TGFB signaling significantly impaired the regenerative response. Intestinal regeneration is characterized by the induction of a fetal-like transcriptional signature during repair. In organoid culture, TGFB1 treatment was necessary and sufficient to induce the fetal-like/regenerative state. Mesenchymal cells were also responsive to TGFB1 and enhanced the regenerative response. Mechanistically, pro-regenerative factors, YAP/TEAD and SOX9, are activated in the epithelium exposed to TGFB1. Finally, pre-treatment with TGFB1 enhanced the ability of primary epithelial cultures to engraft into damaged murine colon, suggesting promise for cellular therapy.
Keywords: Clu; TGFB1; fetal reversion; intestine; macrophage; monocyte; organoid transplantation; regeneration; regenerative medicine; revival stem cell.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests L.C. and M.P.V are listed inventors of a provisional patent application 63392365.
Figures
Update of
-
TGFB1 Induces Fetal Reprogramming and Enhances Intestinal Regeneration.bioRxiv [Preprint]. 2023 Jan 13:2023.01.13.523825. doi: 10.1101/2023.01.13.523825. bioRxiv. 2023. Update in: Cell Stem Cell. 2023 Nov 2;30(11):1520-1537.e8. doi: 10.1016/j.stem.2023.09.015. PMID: 36711781 Free PMC article. Updated. Preprint.
References
-
- Tomic G, Morrissey E, Kozar S, Ben-Moshe S, Hoyle A, Azzarelli R, Kemp R, Chilamakuri CSR, Itzkovitz S, Philpott A, and Winton DJ (2018). Phospho-regulation of ATOH1 Is Required for Plasticity of Secretory Progenitors and Tissue Regeneration. Cell stem cell 23, 436-+. 10.1016/j.stem.2018.07.002. - DOI - PMC - PubMed
-
- Tetteh PW, Basak O, Farin HF, Wiebrands K, Kretzschmar K, Begthel H, van den Born M, Korving J, de Sauvage F, van Es JH, et al. (2016). Replacement of Lost Lgr5-Positive Stem Cells through Plasticity of Their Enterocyte-Lineage Daughters. Cell stem cell 18, 203–213. 10.1016/j.stem.2016.01.001. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P41 RR012408/RR/NCRR NIH HHS/United States
- F31 DK137596/DK/NIDDK NIH HHS/United States
- U01 DK103141/DK/NIDDK NIH HHS/United States
- R01 DK119198/DK/NIDDK NIH HHS/United States
- R01 DK121915/DK/NIDDK NIH HHS/United States
- R01 DK109559/DK/NIDDK NIH HHS/United States
- R01 CA260838/CA/NCI NIH HHS/United States
- R01 DK121166/DK/NIDDK NIH HHS/United States
- R01 DK115806/DK/NIDDK NIH HHS/United States
- R01 DK132885/DK/NIDDK NIH HHS/United States
- R01 DK137806/DK/NIDDK NIH HHS/United States
- P30 CA072720/CA/NCI NIH HHS/United States
- R01 DK126446/DK/NIDDK NIH HHS/United States
- T32 GM139804/GM/NIGMS NIH HHS/United States
- P30 DK034987/DK/NIDDK NIH HHS/United States
- T32 GM135141/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
