

De novo protein identification in mammalian sperm using in situ cryoelectron tomography and AlphaFold2 docking
- PMID: 37865089
- PMCID: PMC10842264
- DOI: 10.1016/j.cell.2023.09.017
De novo protein identification in mammalian sperm using in situ cryoelectron tomography and AlphaFold2 docking
Abstract
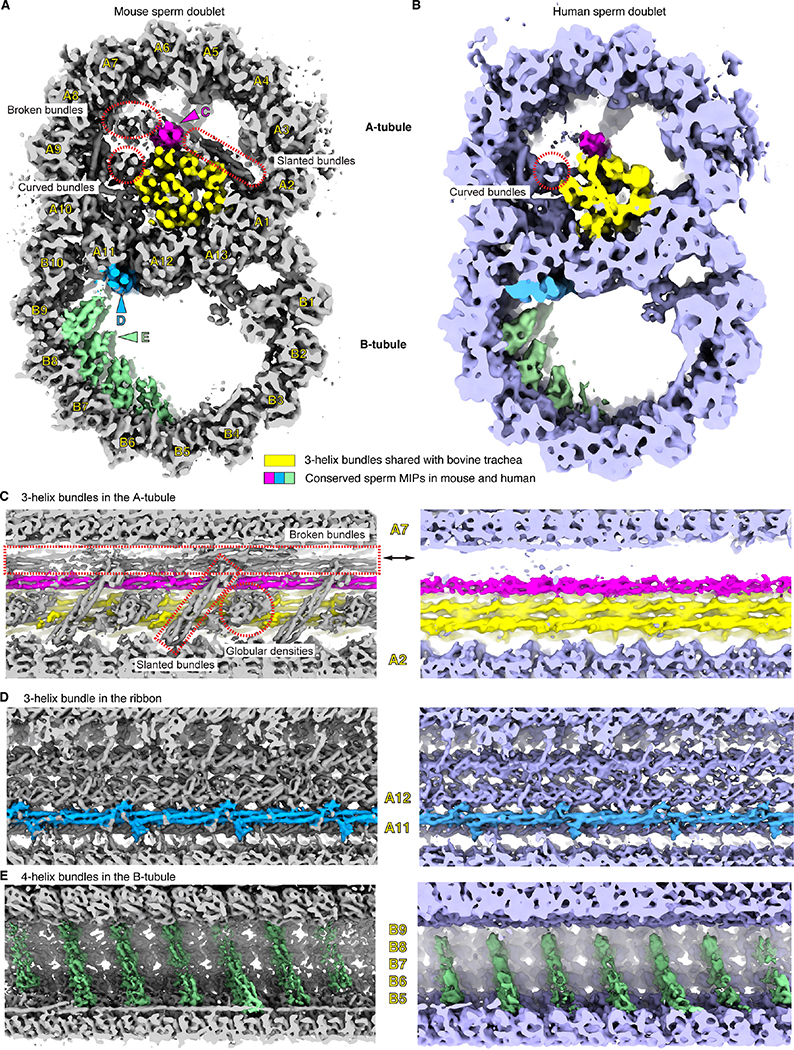
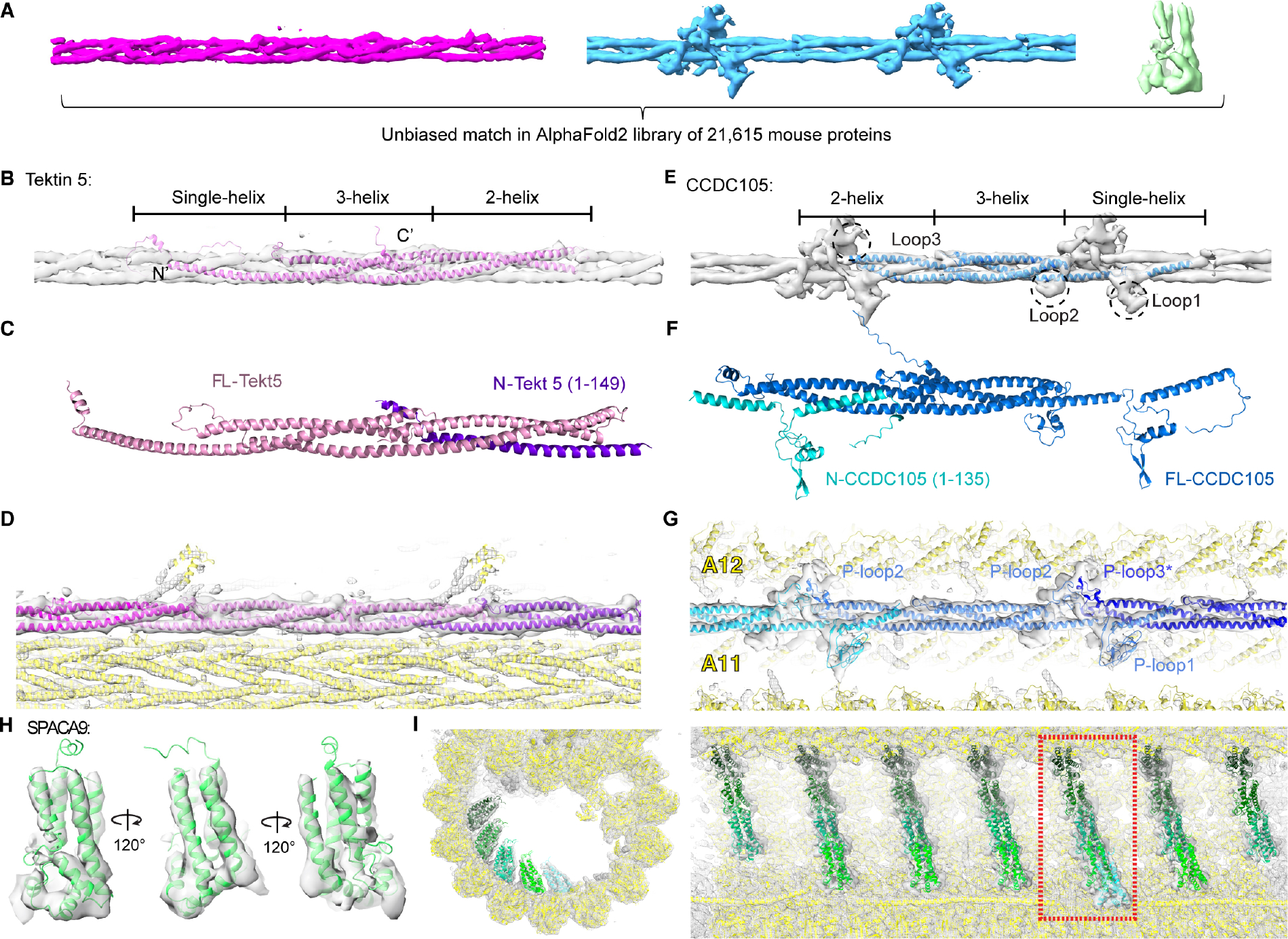
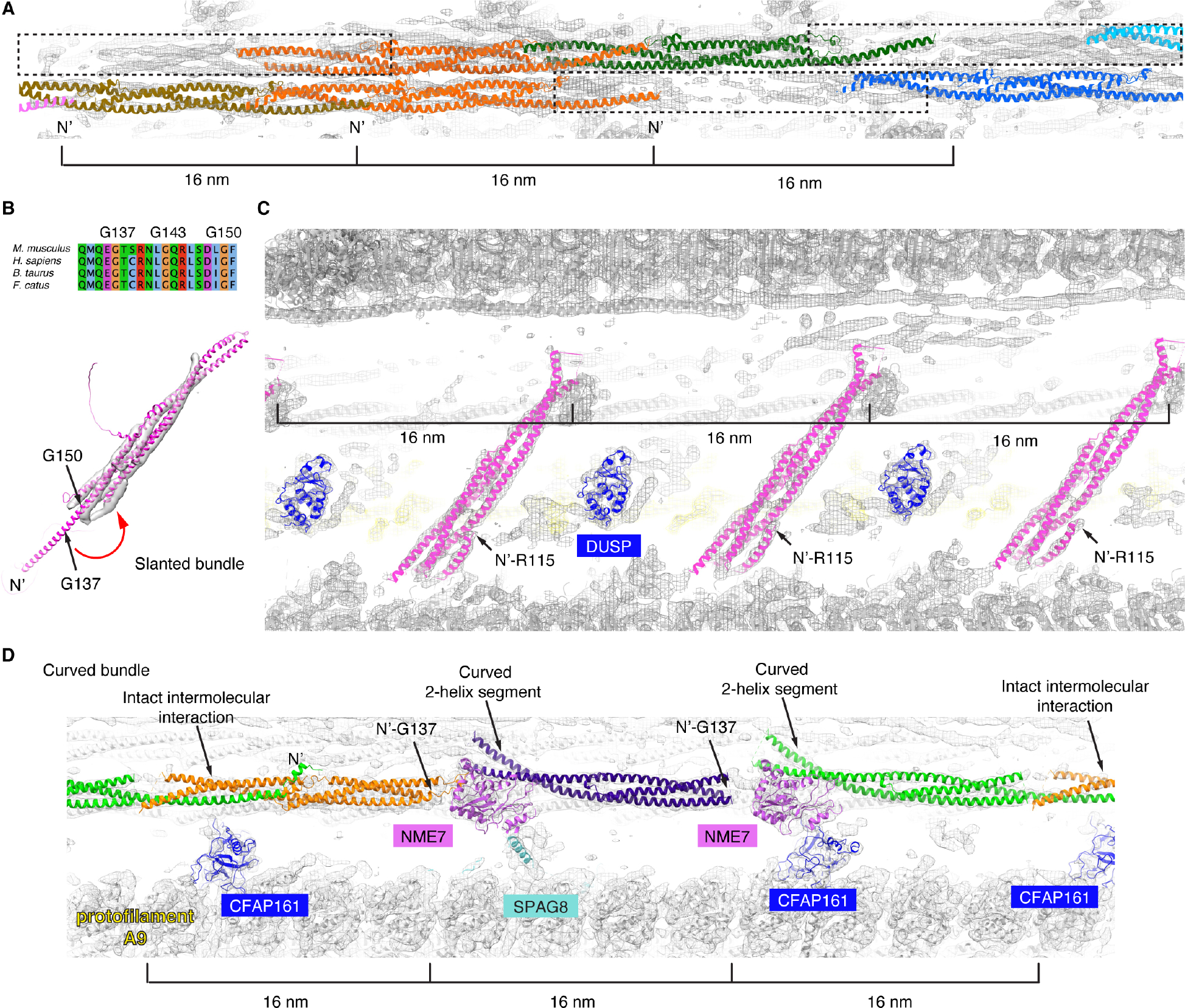
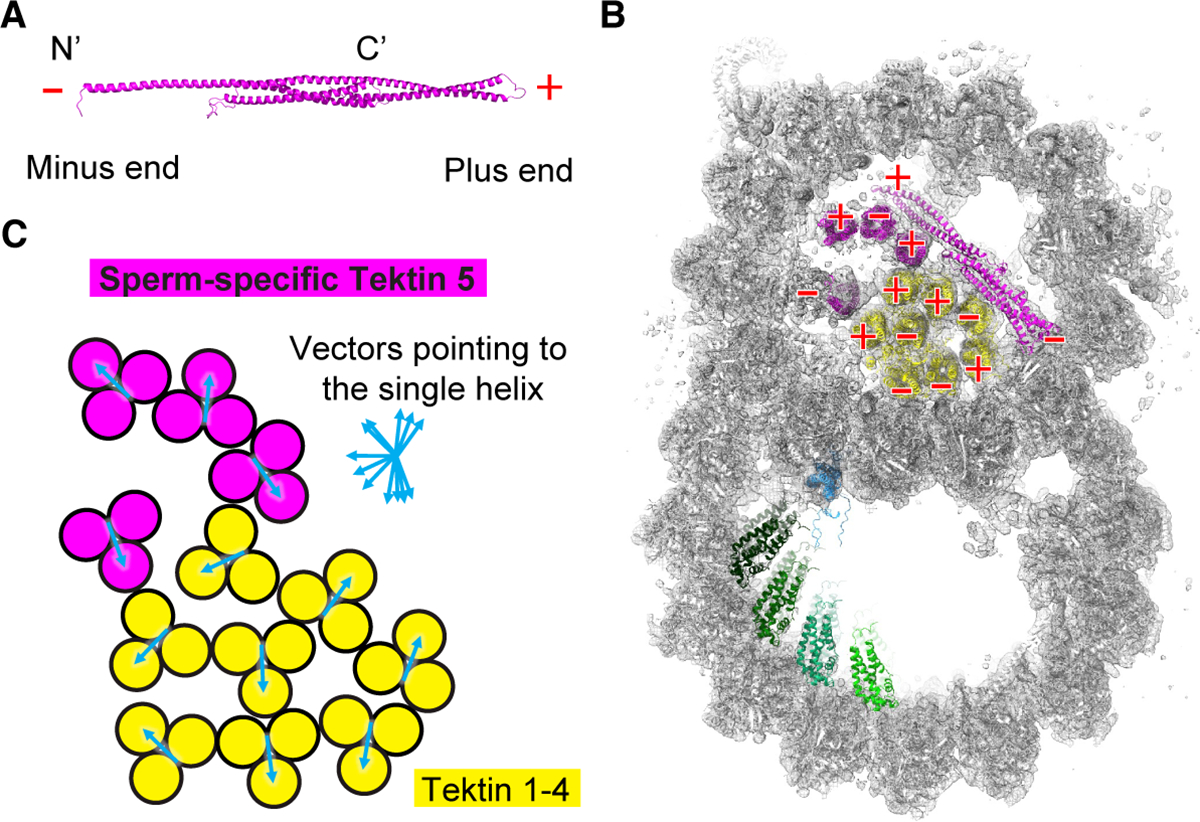
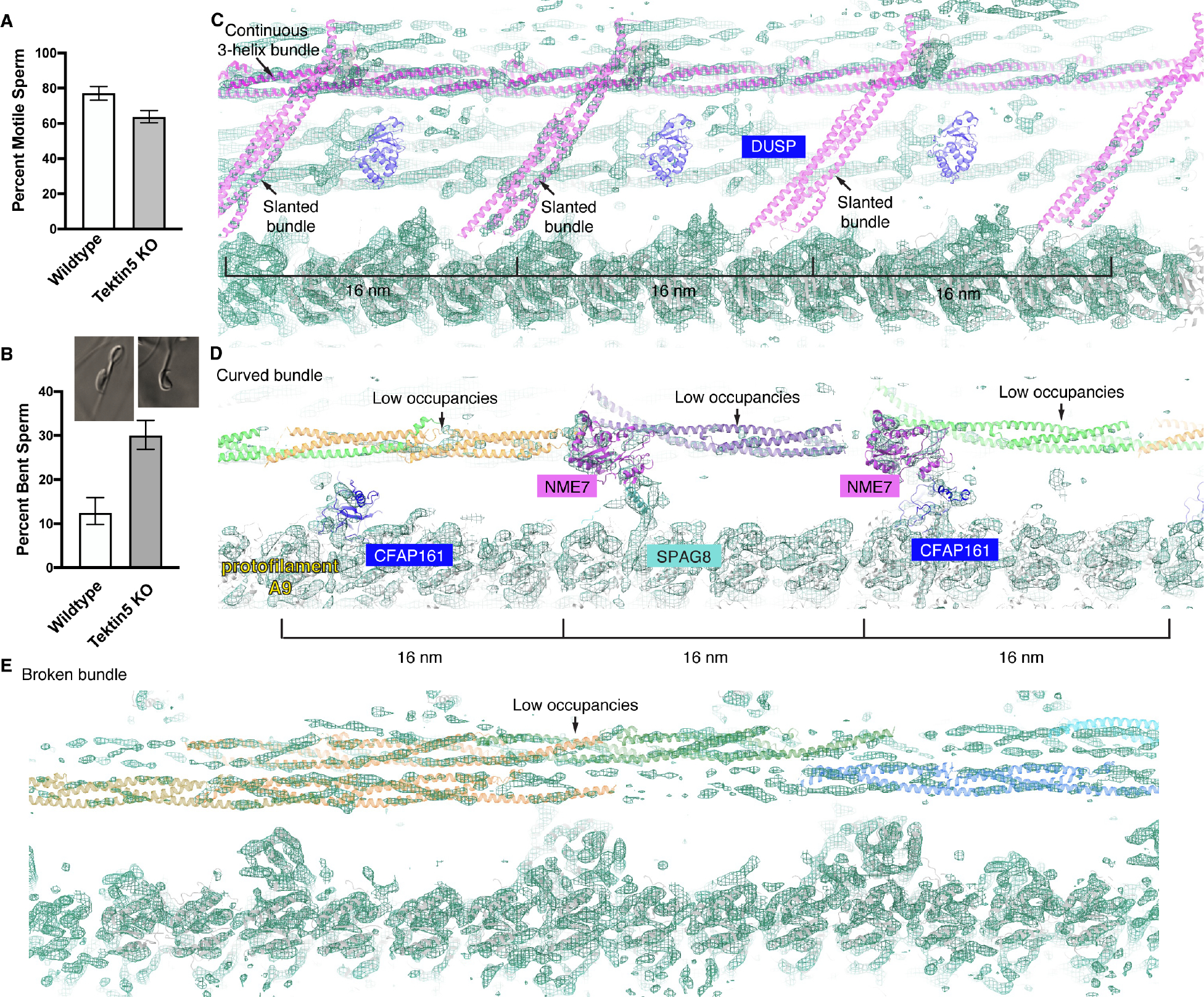
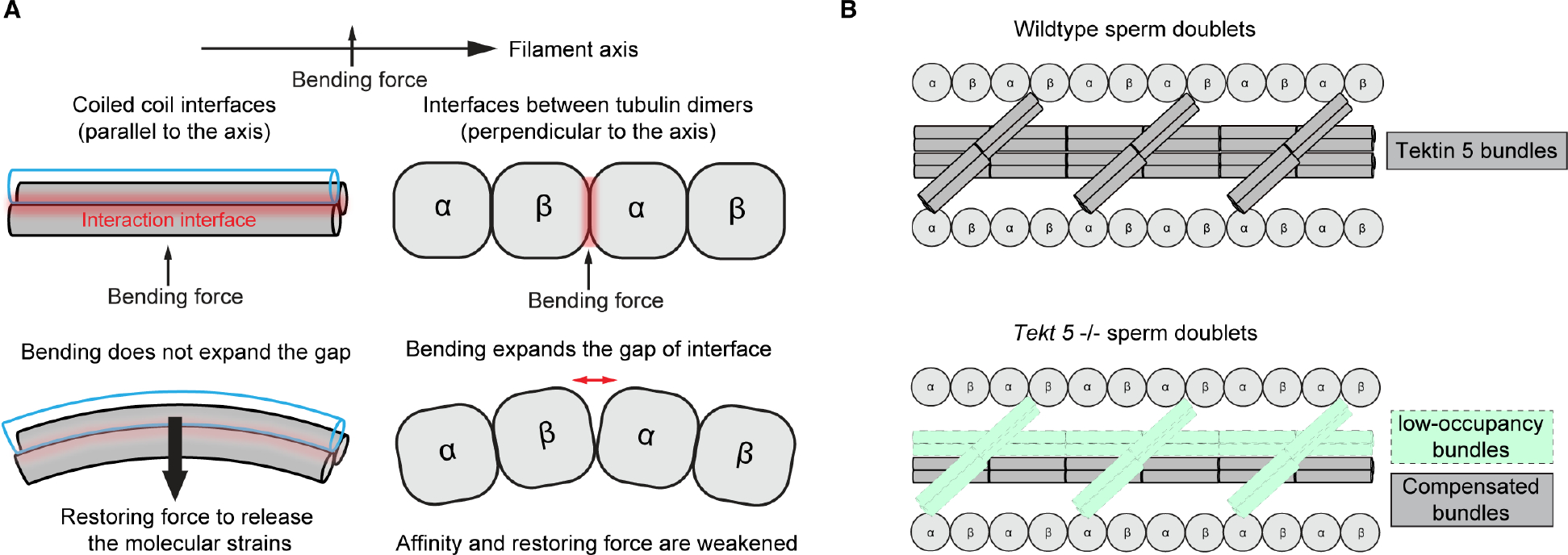
To understand the molecular mechanisms of cellular pathways, contemporary workflows typically require multiple techniques to identify proteins, track their localization, and determine their structures in vitro. Here, we combined cellular cryoelectron tomography (cryo-ET) and AlphaFold2 modeling to address these questions and understand how mammalian sperm are built in situ. Our cellular cryo-ET and subtomogram averaging provided 6.0-Å reconstructions of axonemal microtubule structures. The well-resolved tertiary structures allowed us to unbiasedly match sperm-specific densities with 21,615 AlphaFold2-predicted protein models of the mouse proteome. We identified Tektin 5, CCDC105, and SPACA9 as novel microtubule-associated proteins. These proteins form an extensive interaction network crosslinking the lumen of axonemal doublet microtubules, suggesting their roles in modulating the mechanical properties of the filaments. Indeed, Tekt5 -/- sperm possess more deformed flagella with 180° bends. Together, our studies presented a cellular visual proteomics workflow and shed light on the in vivo functions of Tektin 5.
Keywords: AlphaFold2 modeling; axoneme; cellular cryo-ET; doublet; microtubule; microtubule inner protein; sperm; visual proteomics.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests We declare that one or more authors have a competing interest as defined by Nature Portfolio. The Krogan Laboratory has received research support from Vir Biotechnology, F. Hoffmann-La Roche, and Rezo Therapeutics. N.J.K. has previously held financially compensated consulting agreements with the Icahn School of Medicine at Mount Sinai, New York, and Twist Bioscience Corp. He currently has financially compensated consulting agreements with Maze Therapeutics, Interline Therapeutics, Rezo Therapeutics, and GEn1E Lifesciences, Inc. He is on the Board of Directors of Rezo Therapeutics and is a shareholder in Tenaya Therapeutics, Maze Therapeutics, Rezo Therapeutics, and Interline Therapeutics.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
