

Mechanopathology of biofilm-like Mycobacterium tuberculosis cords
- PMID: 37865090
- PMCID: PMC10642369
- DOI: 10.1016/j.cell.2023.09.016
Mechanopathology of biofilm-like Mycobacterium tuberculosis cords
Abstract
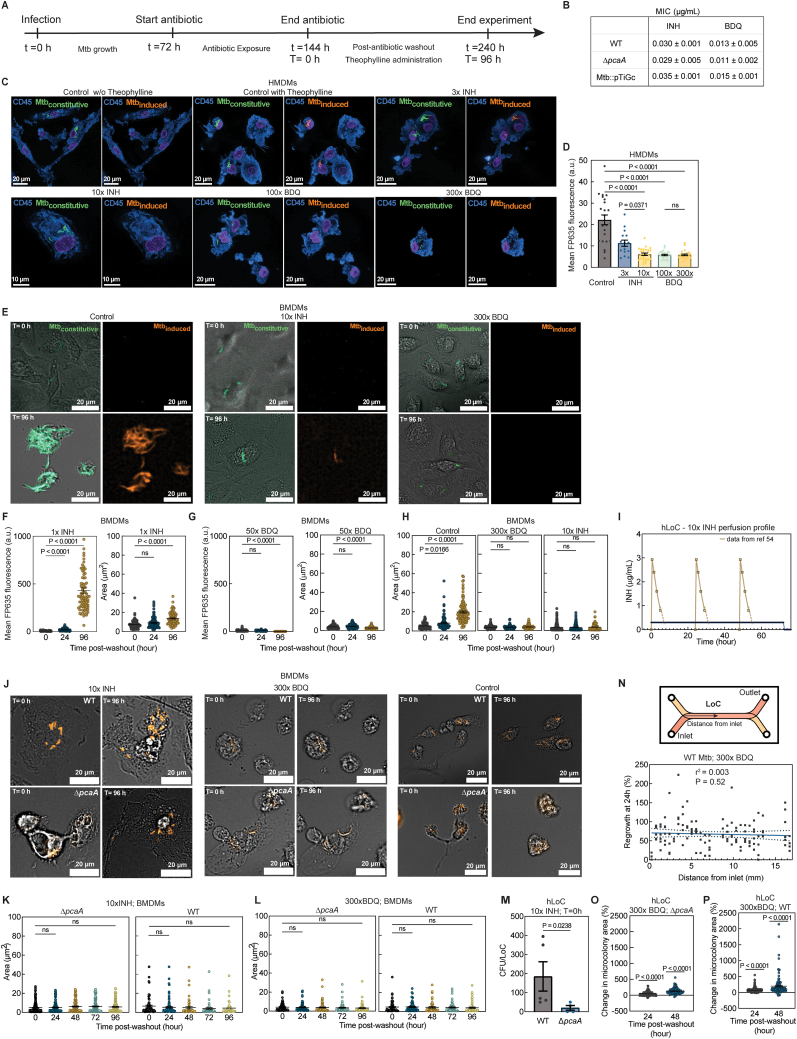
Mycobacterium tuberculosis (Mtb) cultured axenically without detergent forms biofilm-like cords, a clinical identifier of virulence. In lung-on-chip (LoC) and mouse models, cords in alveolar cells contribute to suppression of innate immune signaling via nuclear compression. Thereafter, extracellular cords cause contact-dependent phagocyte death but grow intercellularly between epithelial cells. The absence of these mechanopathological mechanisms explains the greater proportion of alveolar lesions with increased immune infiltration and dissemination defects in cording-deficient Mtb infections. Compression of Mtb lipid monolayers induces a phase transition that enables mechanical energy storage. Agent-based simulations demonstrate that the increased energy storage capacity is sufficient for the formation of cords that maintain structural integrity despite mechanical perturbation. Bacteria in cords remain translationally active despite antibiotic exposure and regrow rapidly upon cessation of treatment. This study provides a conceptual framework for the biophysics and function in tuberculosis infection and therapy of cord architectures independent of mechanisms ascribed to single bacteria.
Keywords: Mycobacterium tuberculosis; agent-based model; antibiotic therapy; biofilms; cords; lung-on-chip; mechanobiology; mycomembrane; serial block scanning face electron microscopy.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures













Comment in
-
Mechanobiology: How pathogens use mechanics to modulate host interactions.Cell. 2023 Nov 9;186(23):4994-4995. doi: 10.1016/j.cell.2023.09.018. Cell. 2023. PMID: 37949055
References
-
- Ojha A.K., Baughn A.D., Sambandan D., Hsu T., Trivelli X., Guerardel Y., Alahari A., Kremer L., Jacobs W.R., Hatfull G.F. Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria. Mol. Microbiol. 2008;69:164–174. doi: 10.1111/j.1365-2958.2008.06274.x. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
