

Wnt/β-catenin and NFκB signaling synergize to trigger growth factor-free regeneration of adult primary human hepatocytes
- PMID: 37870288
- PMCID: PMC11095891
- DOI: 10.1097/HEP.0000000000000648
Wnt/β-catenin and NFκB signaling synergize to trigger growth factor-free regeneration of adult primary human hepatocytes
Abstract
Background and aims: The liver has a remarkable capacity to regenerate, which is sustained by the ability of hepatocytes to act as facultative stem cells that, while normally quiescent, re-enter the cell cycle after injury. Growth factor signaling is indispensable in rodents, whereas Wnt/β-catenin is not required for effective tissue repair. However, the molecular networks that control human liver regeneration remain unclear.
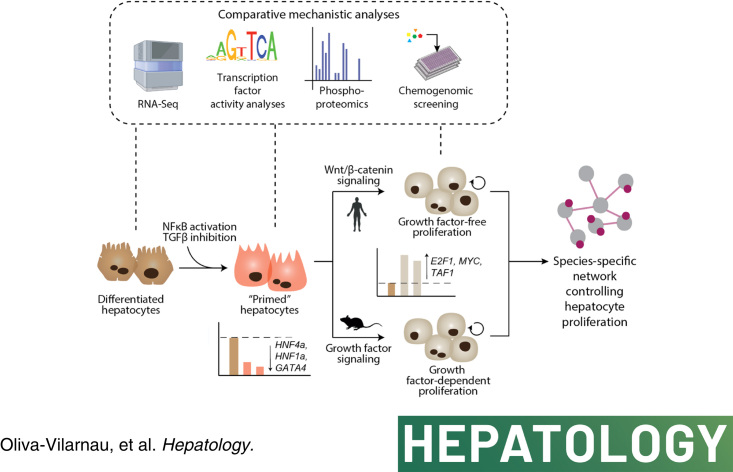
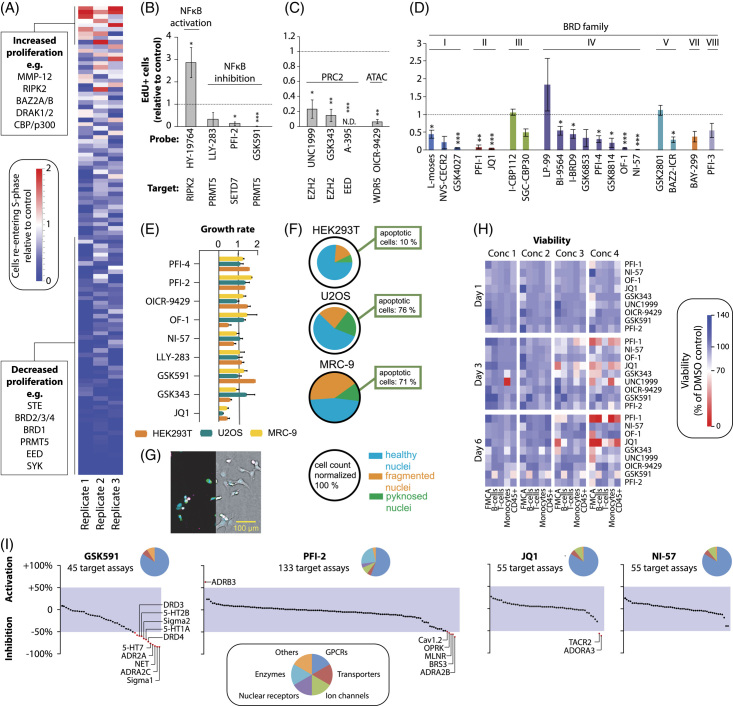
Methods: Organotypic 3D spheroid cultures of primary human or murine hepatocytes were used to identify the signaling network underlying cell cycle re-entry. Furthermore, we performed chemogenomic screening of a library enriched for epigenetic regulators and modulators of immune function to determine the importance of epigenomic control for human hepatocyte regeneration.
Results: Our results showed that, unlike in rodents, activation of Wnt/β-catenin signaling is the major mitogenic cue for adult primary human hepatocytes. Furthermore, we identified TGFβ inhibition and inflammatory signaling through NF-κB as essential steps for the quiescent-to-regenerative switch that allows Wnt/β-catenin-induced proliferation of human cells. In contrast, growth factors, but not Wnt/β-catenin signaling, triggered hyperplasia in murine hepatocytes. High-throughput screening in a human model confirmed the relevance of NFκB and revealed the critical roles of polycomb repressive complex 2, as well as of the bromodomain families I, II, and IV.
Conclusions: This study revealed a network of NFκB, TGFβ, and Wnt/β-catenin that controls human hepatocyte regeneration in the absence of exogenous growth factors, identified novel regulators of hepatocyte proliferation, and highlighted the potential of organotypic culture systems for chemogenomic interrogation of complex physiological processes.
Copyright © 2023 The Author(s). Published by Wolters Kluwer Health, Inc.
Conflict of interest statement
Volker M. Lauschke is employed by and owns stock in HepaPredict. He owns stock in PersoMedix. The remaining authors have no conflicts to report.
Figures





Comment in
-
Hepatocytes of mice and men: Different regenerative signals?Hepatology. 2024 Jun 1;79(6):1246-1248. doi: 10.1097/HEP.0000000000000693. Epub 2023 Nov 16. Hepatology. 2024. PMID: 37972957 No abstract available.
References
-
- Miyaoka Y, Ebato K, Kato H, Arakawa S, Shimizu S, Miyajima A. Hypertrophy and unconventional cell division of hepatocytes underlie liver regeneration. Curr Biol. 2012;22:1166–1175. - PubMed
-
- MacDonald RA. Lifespan of liver cells: Autoradiographic study using tritiated thymidine in normal, cirrhotic, and partially hepatectomized rats. Arch Intern Med. 1961;107:335–343. - PubMed
-
- Wang MJ, Chen F, Li JX, Liu CC, Zhang HB, Xia Y, et al. Reversal of hepatocyte senescence after continuous in vivo cell proliferation. Hepatology. 2014;60:349–361. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
