

A mechanism-based theory of cellular and tissue plasticity
- PMID: 37871208
- PMCID: PMC10622945
- DOI: 10.1073/pnas.2305375120
A mechanism-based theory of cellular and tissue plasticity
Abstract
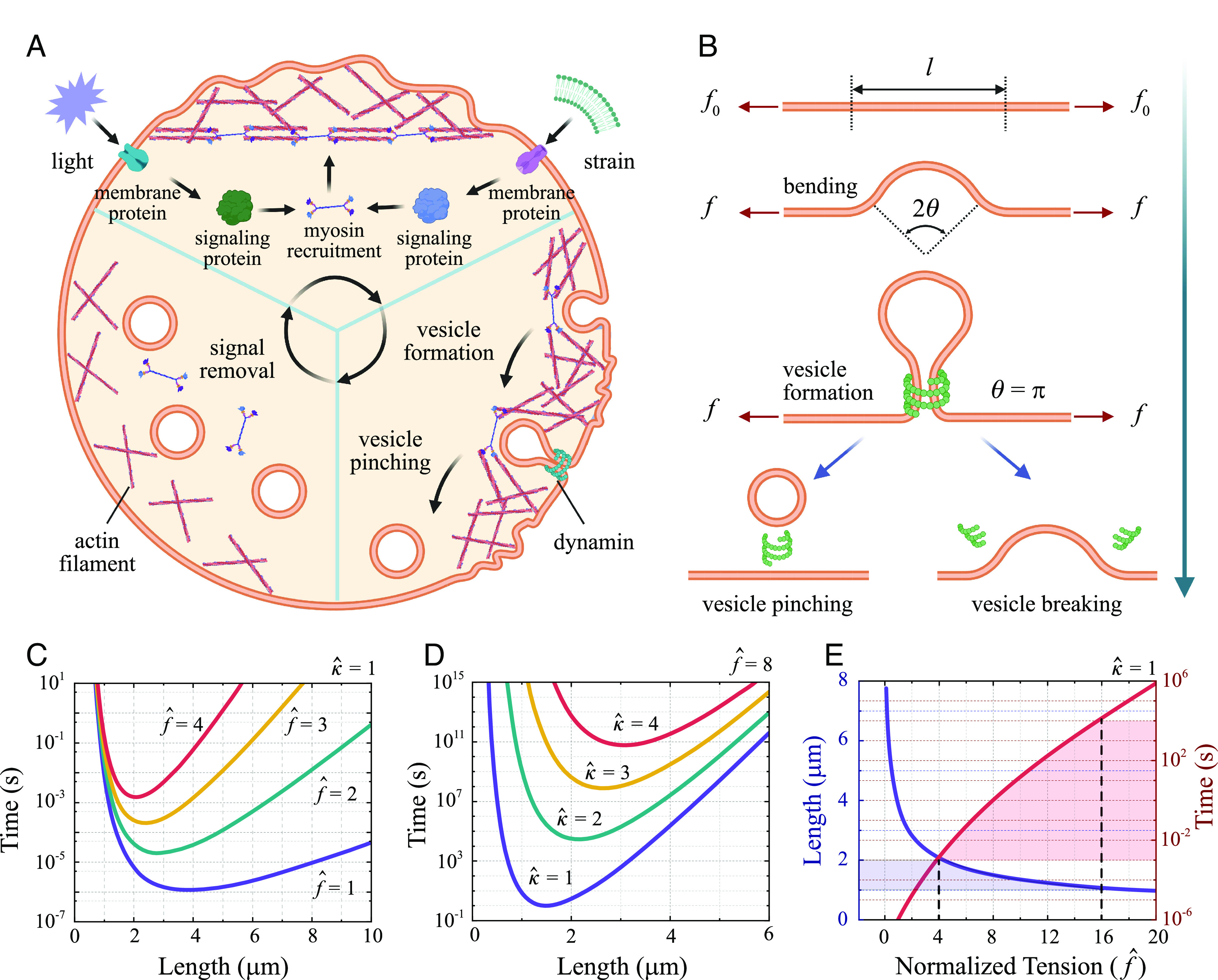
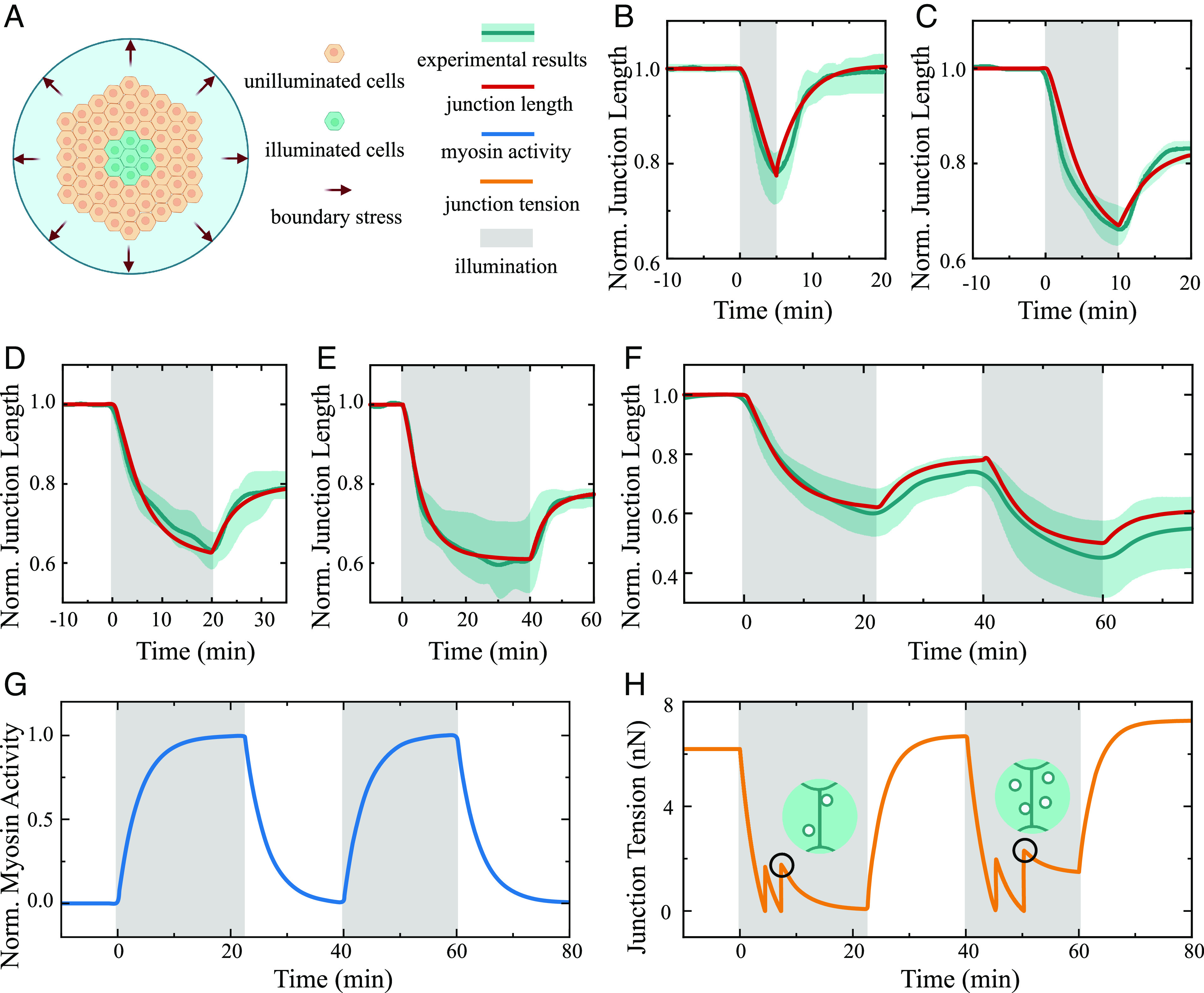
Plastic deformation in cells and tissues has been found to play crucial roles in collective cell migration, cancer metastasis, and morphogenesis. However, the fundamental question of how plasticity is initiated in individual cells and then propagates within the tissue remains elusive. Here, we develop a mechanism-based theory of cellular and tissue plasticity that accounts for all key processes involved, including the activation and development of active contraction at different scales as well as the formation of endocytic vesicles on cell junctions and show that this theory achieves quantitative agreement with all existing experiments. Specifically, it reveals that, in response to optical or mechanical stimuli, the myosin contraction and thermal fluctuation-assisted formation and pinching of endocytic vesicles could lead to permanent shortening of cell junctions and that such plastic constriction can stretch neighboring cells and trigger their active contraction through mechanochemical feedbacks and eventually their plastic deformations as well. Our theory predicts that endocytic vesicles with a size around 1 to 2 µm will most likely be formed and a higher irreversible shortening of cell junctions could be achieved if a long stimulation is split into multiple short ones, all in quantitative agreement with experiments. Our analysis also shows that constriction of cells in tissue can undergo elastic/unratcheted to plastic/ratcheted transition as the magnitude and duration of active contraction increases, ultimately resulting in the propagation of plastic deformation waves within the monolayer with a constant speed which again is consistent with experimental observations.
Keywords: active contraction; cell plasticity; endocytosis; morphogenesis; tissue wave.
Conflict of interest statement
The authors declare no competing interest.
Figures



Similar articles
-
Microfilaments in cellular and developmental processes.Science. 1971 Jan 15;171(3967):135-43. doi: 10.1126/science.171.3967.135. Science. 1971. PMID: 5538822
-
ERK-Mediated Mechanochemical Waves Direct Collective Cell Polarization.Dev Cell. 2020 Jun 22;53(6):646-660.e8. doi: 10.1016/j.devcel.2020.05.011. Epub 2020 Jun 3. Dev Cell. 2020. PMID: 32497487
-
Apical constriction: themes and variations on a cellular mechanism driving morphogenesis.Development. 2014 May;141(10):1987-98. doi: 10.1242/dev.102228. Development. 2014. PMID: 24803648 Free PMC article. Review.
-
[Regulation of intercellular adhesion during epithelial morphogenesis].Biol Aujourdhui. 2012;206(3):219-36. doi: 10.1051/jbio/2012021. Epub 2012 Nov 22. Biol Aujourdhui. 2012. PMID: 23171844 Review. French.
-
Inverse tissue mechanics of cell monolayer expansion.PLoS Comput Biol. 2018 Mar 1;14(3):e1006029. doi: 10.1371/journal.pcbi.1006029. eCollection 2018 Mar. PLoS Comput Biol. 2018. PMID: 29494578 Free PMC article.
Cited by
-
Atomic Force Microscopy for the Study of Cell Mechanics in Pharmaceutics.Pharmaceutics. 2024 May 29;16(6):733. doi: 10.3390/pharmaceutics16060733. Pharmaceutics. 2024. PMID: 38931854 Free PMC article. Review.
-
An actin bracket-induced elastoplastic transition determines epithelial folding irreversibility.Nat Commun. 2024 Dec 12;15(1):10476. doi: 10.1038/s41467-024-54906-7. Nat Commun. 2024. PMID: 39668169 Free PMC article.
References
-
- Gilmour D., Rembold M., Leptin M., From morphogen to morphogenesis and back. Nature 541, 311–320 (2017). - PubMed
-
- Bonakdar N., et al. , Mechanical plasticity of cells. Nat. Mater. 15, 1090–1094 (2016). - PubMed
-
- Jain S., Ladoux B., Mege R. M., Mechanical plasticity in collective cell migration. Curr. Opin. Cell Biol. 72, 54–62 (2021). - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
