

Acylation of glycerolipids in mycobacteria
- PMID: 37872138
- PMCID: PMC10593935
- DOI: 10.1038/s41467-023-42478-x
Acylation of glycerolipids in mycobacteria
Abstract
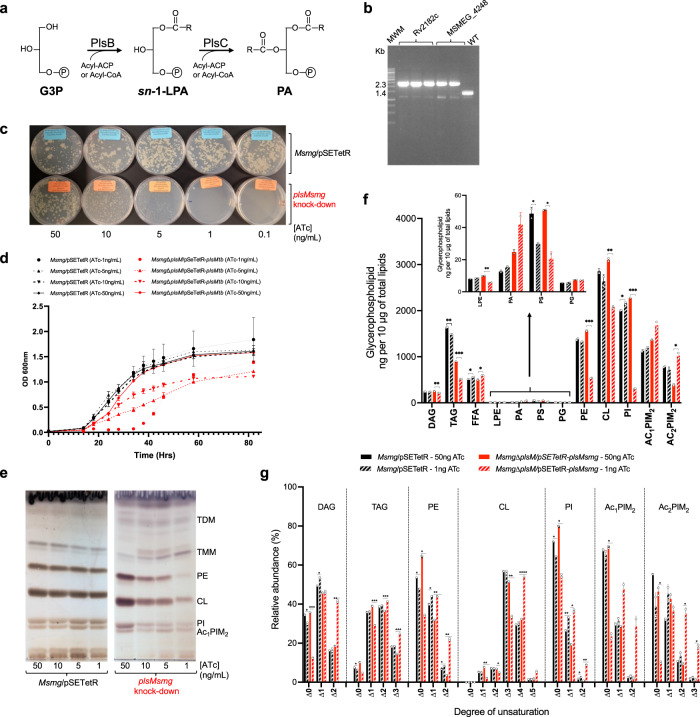
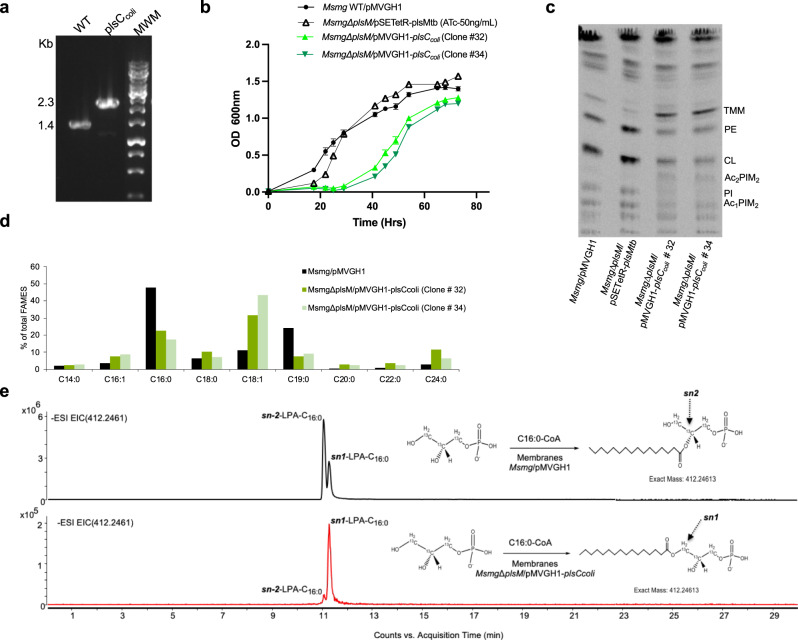
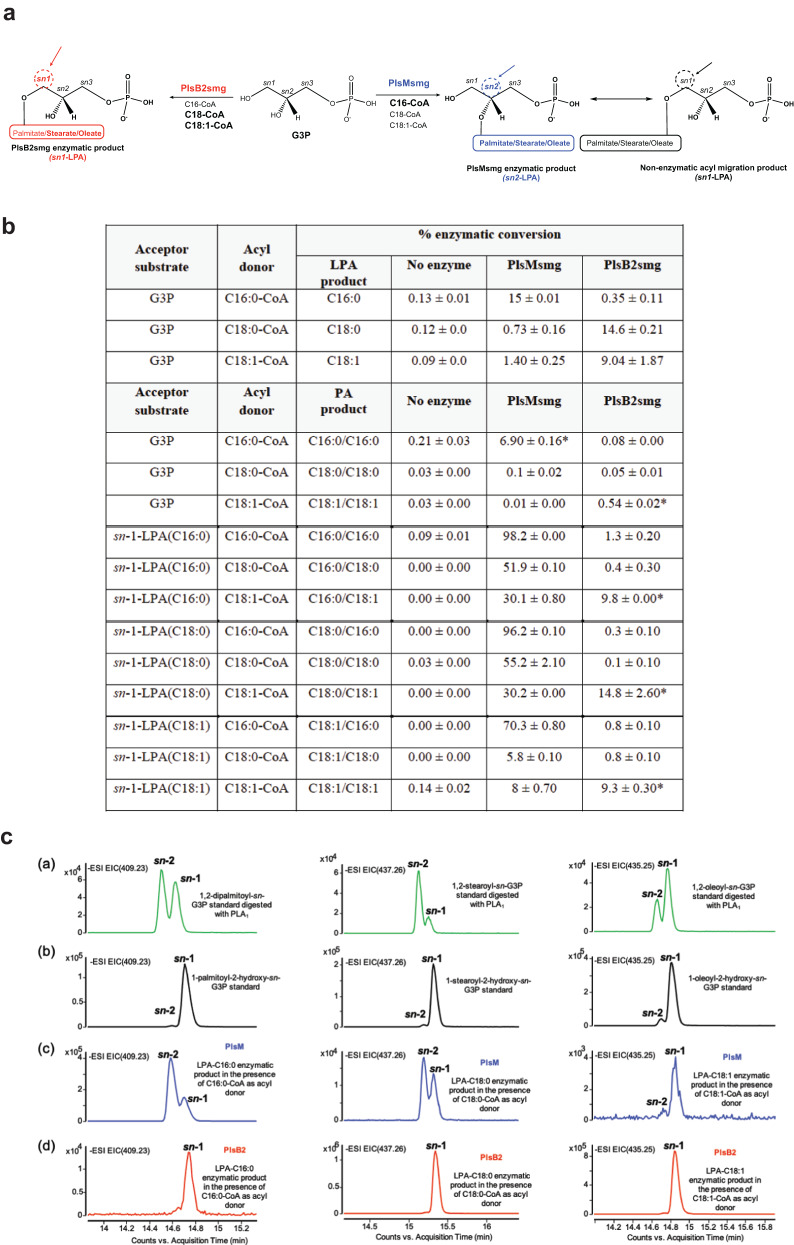
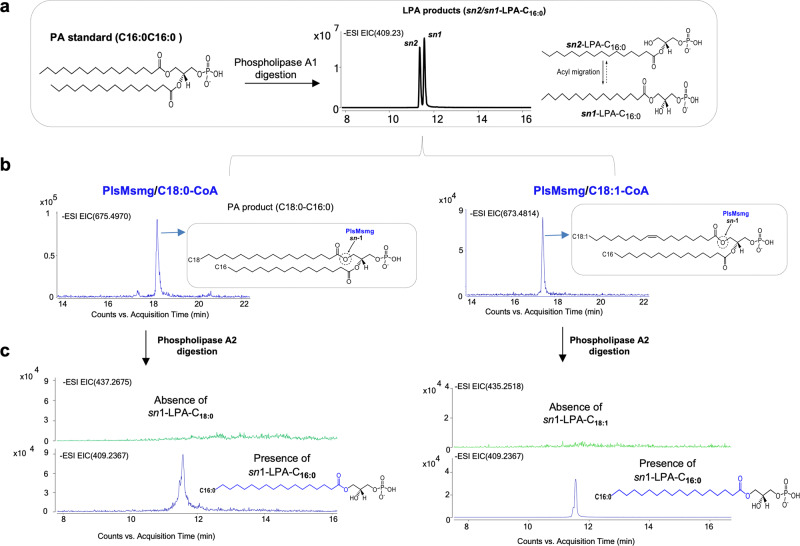
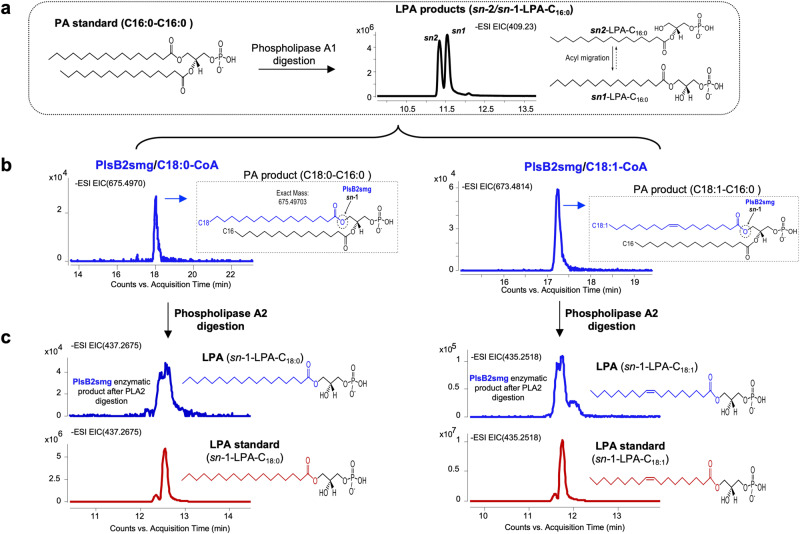
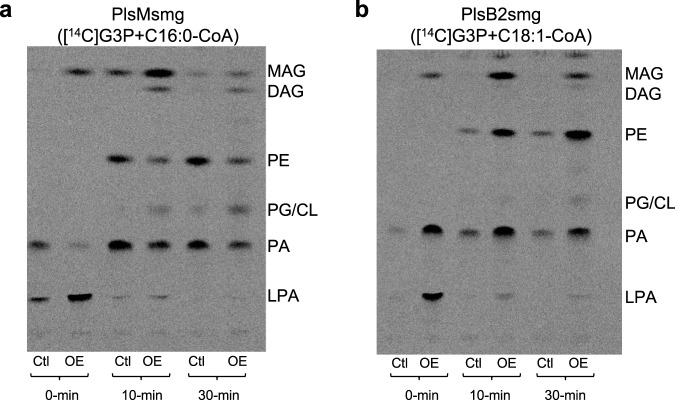
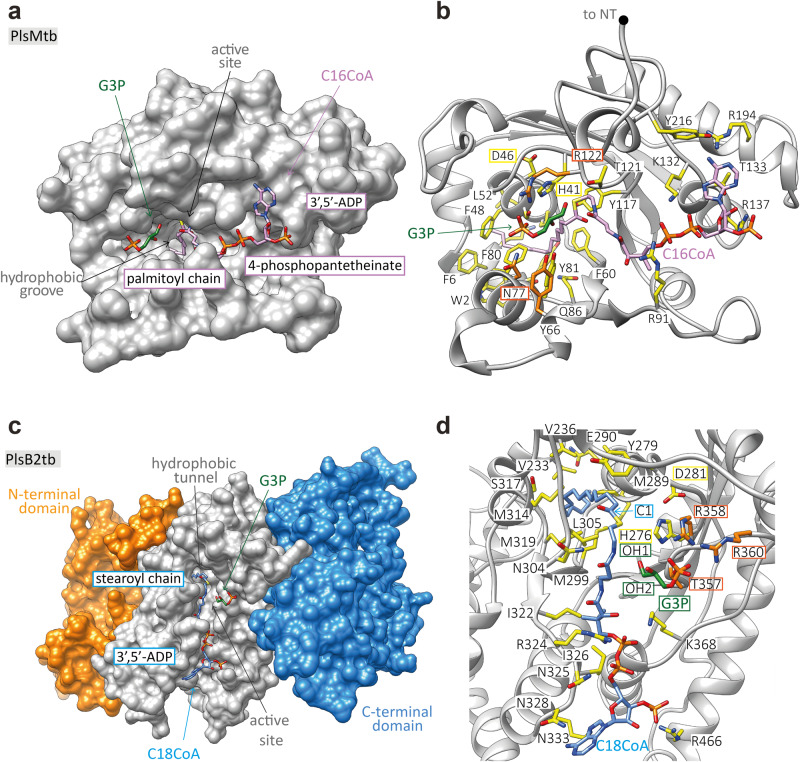
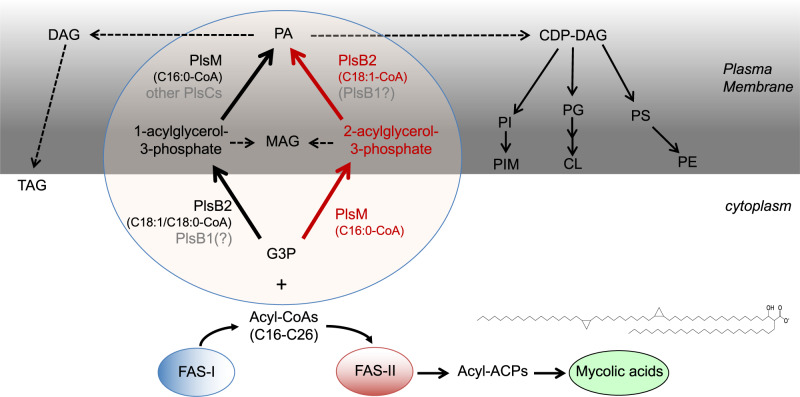
We report on the existence of two phosphatidic acid biosynthetic pathways in mycobacteria, a classical one wherein the acylation of the sn-1 position of glycerol-3-phosphate (G3P) precedes that of sn-2 and another wherein acylations proceed in the reverse order. Two unique acyltransferases, PlsM and PlsB2, participate in both pathways and hold the key to the unusual positional distribution of acyl chains typifying mycobacterial glycerolipids wherein unsaturated substituents principally esterify position sn-1 and palmitoyl principally occupies position sn-2. While PlsM selectively transfers a palmitoyl chain to the sn-2 position of G3P and sn-1-lysophosphatidic acid (LPA), PlsB2 preferentially transfers a stearoyl or oleoyl chain to the sn-1 position of G3P and an oleyl chain to sn-2-LPA. PlsM is the first example of an sn-2 G3P acyltransferase outside the plant kingdom and PlsB2 the first example of a 2-acyl-G3P acyltransferase. Both enzymes are unique in their ability to catalyze acyl transfer to both G3P and LPA.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Goren, M. B. in The mycobacteria. A sourcebook, 1st edn, Vol. 1 (eds. Kubica, G. P. & Wayne, L. G.) 379–415 (Marcel Dekker, 1984).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
