

This is a preprint.
Commensal myeloid crosstalk in neonatal skin regulates cutaneous type 17 inflammation
- PMID: 37873143
- PMCID: PMC10592812
- DOI: 10.1101/2023.09.29.560039
Commensal myeloid crosstalk in neonatal skin regulates cutaneous type 17 inflammation
Abstract
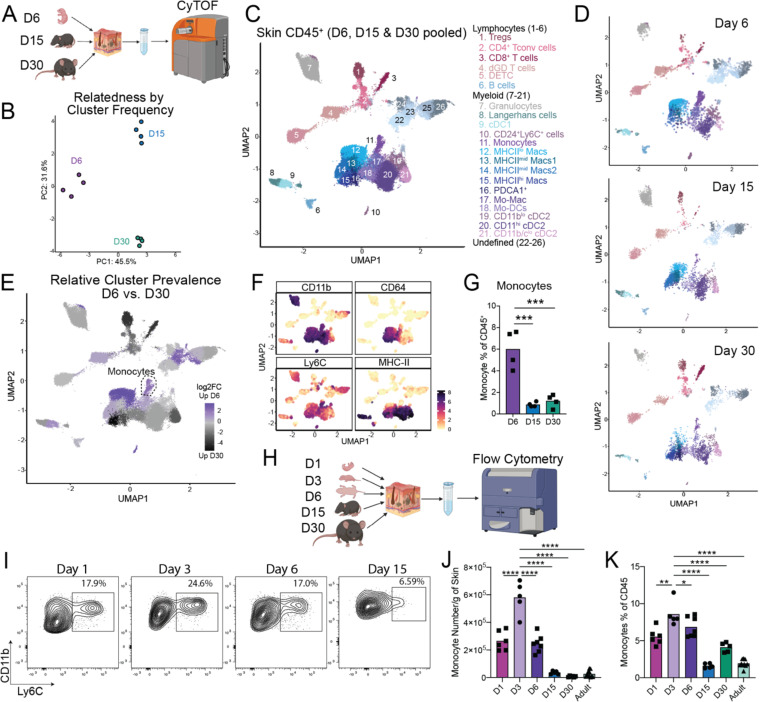
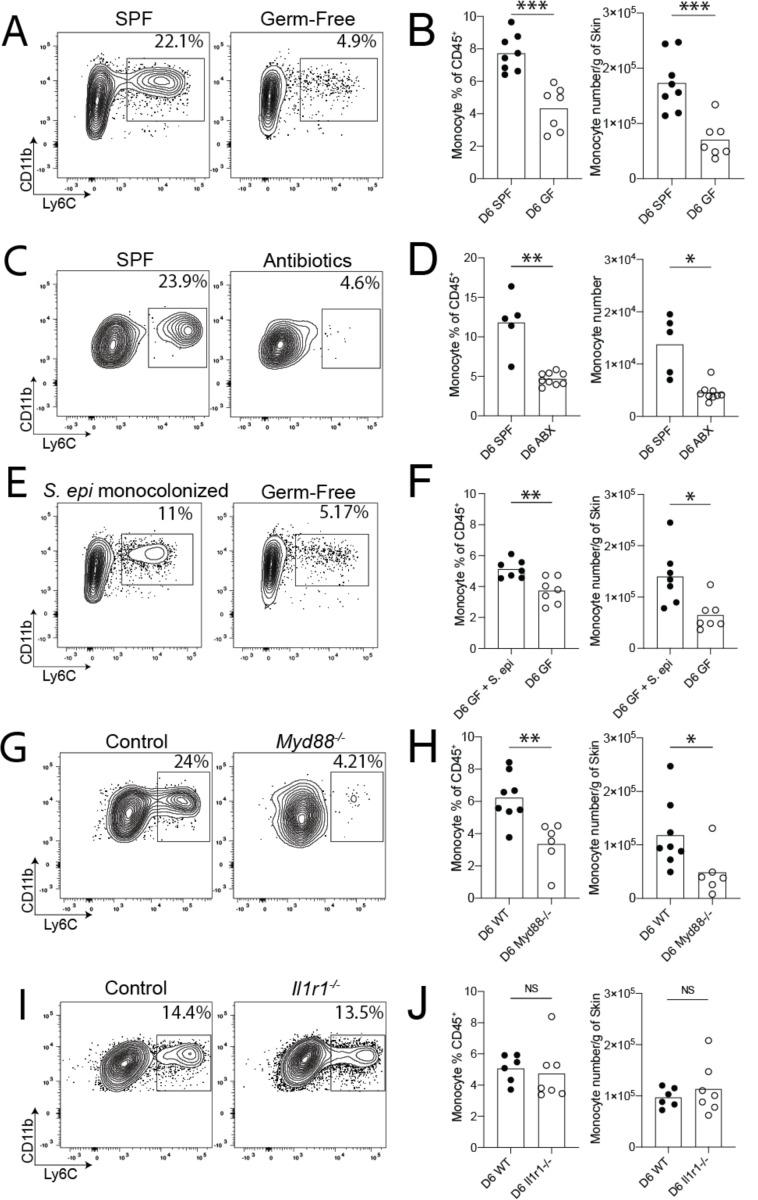
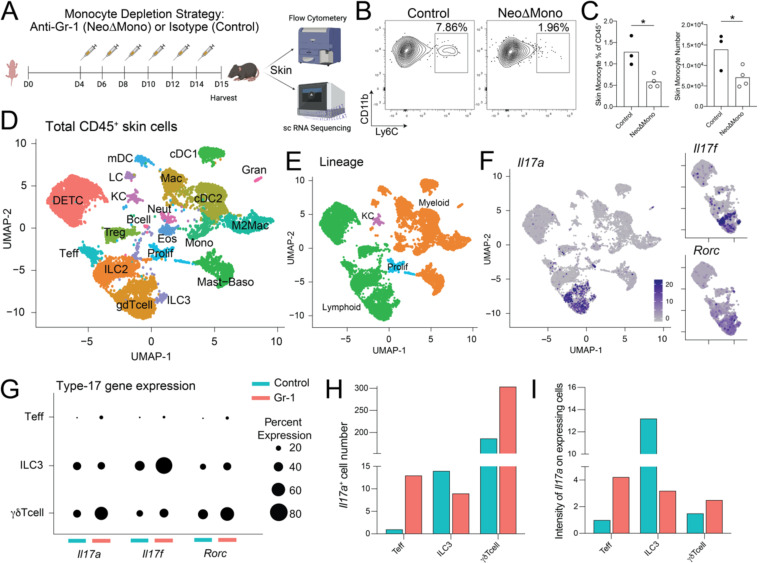
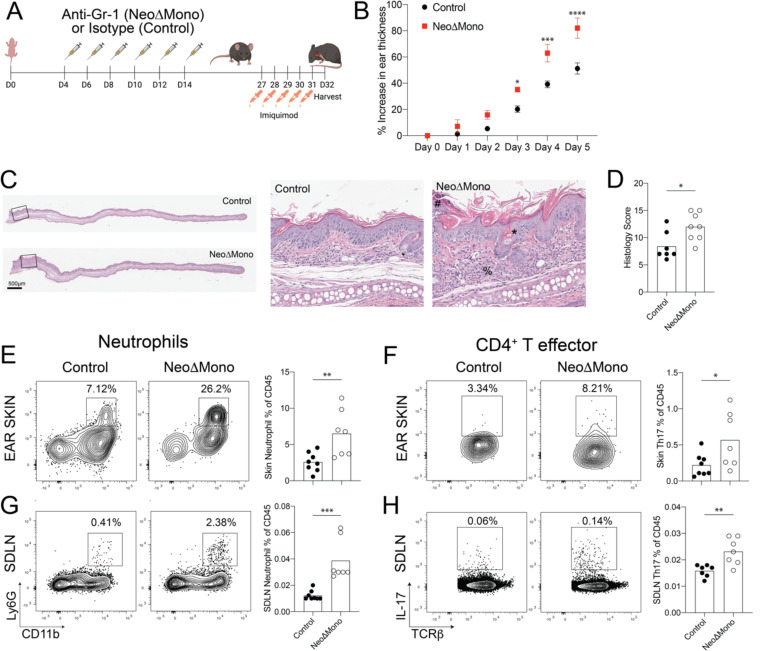
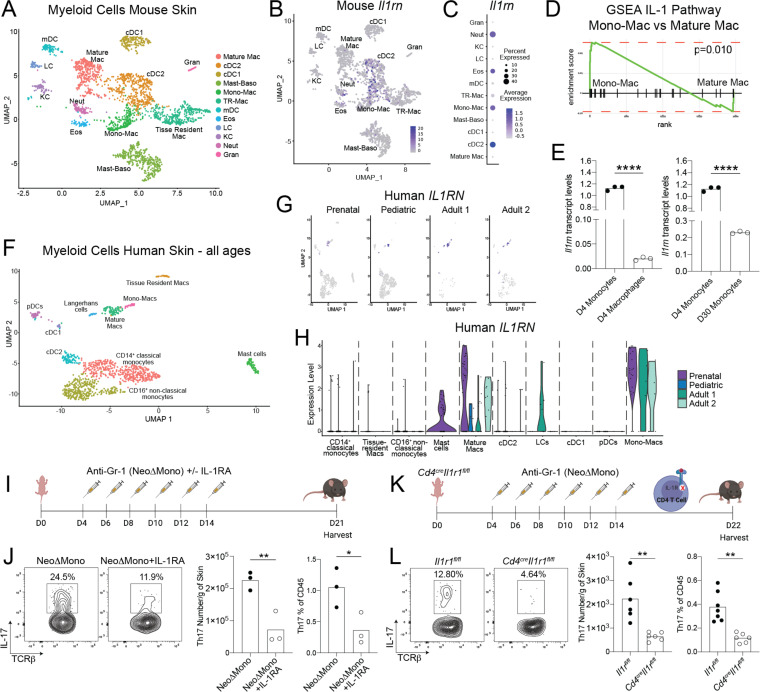
Early life microbe-immune interactions at barrier surfaces have lasting impacts on the trajectory towards health versus disease. Monocytes, macrophages and dendritic cells are primary sentinels in barrier tissues, yet the salient contributions of commensal-myeloid crosstalk during tissue development remain poorly understood. Here, we identify that commensal microbes facilitate accumulation of a population of monocytes in neonatal skin. Transient postnatal depletion of these monocytes resulted in heightened IL-17A production by skin T cells, which was particularly sustained among CD4+ T cells and sufficient to exacerbate inflammatory skin pathologies. Neonatal skin monocytes were enriched in expression of negative regulators of the IL-1 pathway. Functional in vivo experiments confirmed a key role for excessive IL-1R1 signaling in T cells as contributing to the dysregulated type 17 response in neonatal monocyte-depleted mice. Thus, a commensal-driven wave of monocytes into neonatal skin critically facilitates immune homeostasis in this prominent barrier tissue.
Conflict of interest statement
T.C.S is on the scientific advisory board of Concerto Biosciences. M.H.S. is founder, shareholder and board member of Teiko.bio, has received a speaking honorarium from Fluidigm Inc., Kumquat Bio, and Arsenal Bio, has been a paid consultant for Five Prime, Ono, January, Earli, Astellas, and Indaptus, and has received research funding from Roche/Genentech, Pfizer, Valitor, and Bristol Myers Squibb.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials