

This is a preprint.
A developmentally defined population of neurons in the lateral septum controls responses to aversive stimuli
- PMID: 37873286
- PMCID: PMC10592641
- DOI: 10.1101/2023.09.24.559205
A developmentally defined population of neurons in the lateral septum controls responses to aversive stimuli
Abstract
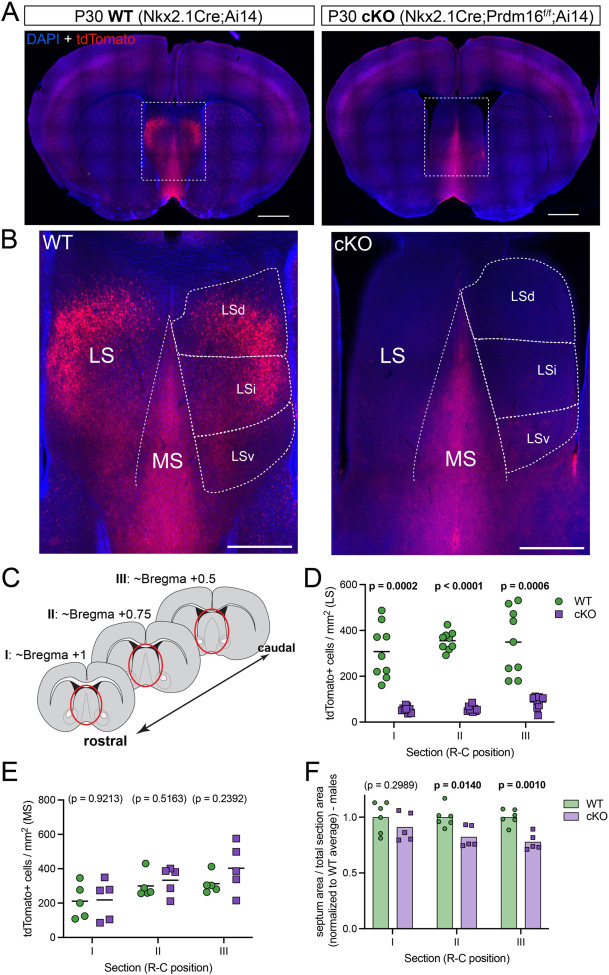
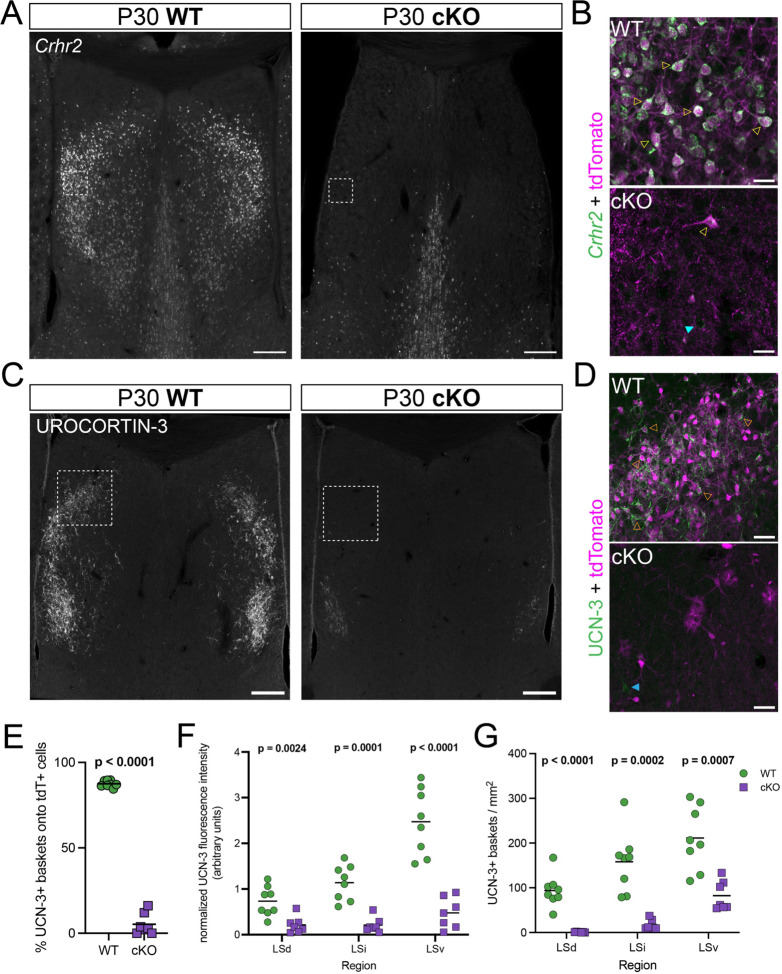
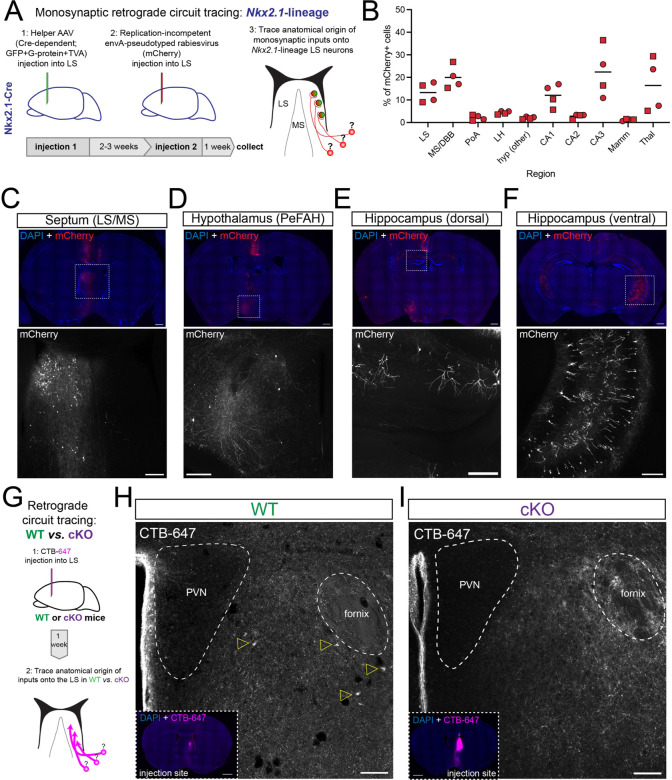
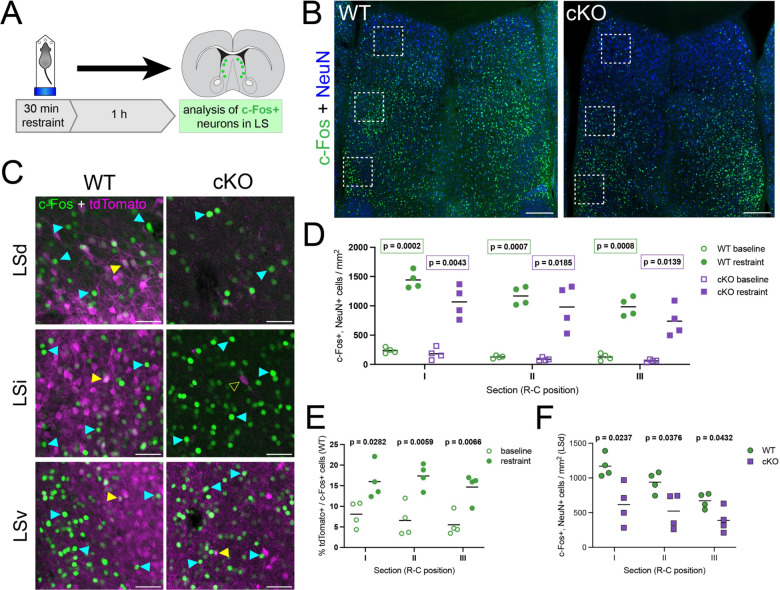
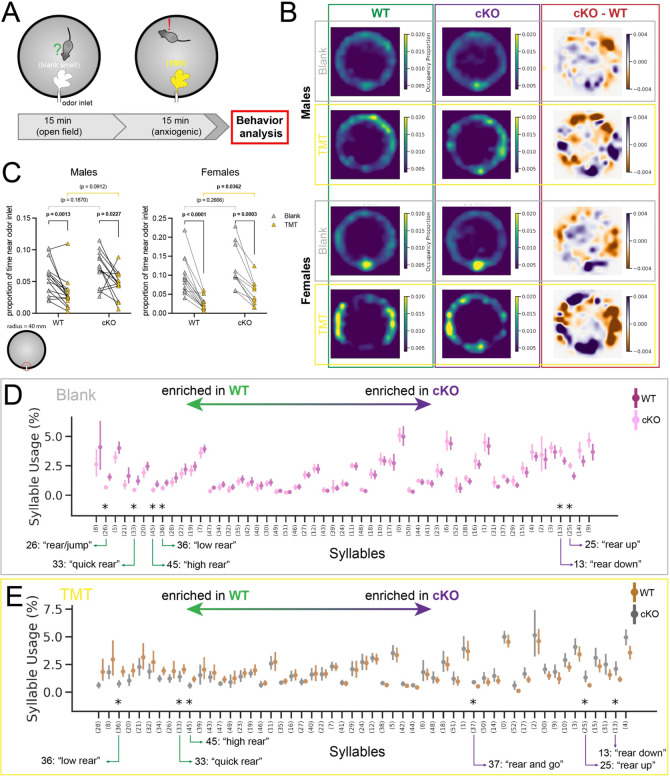
When interacting with their environment, animals must balance exploratory and defensive behavior to evaluate and respond to potential threats. The lateral septum (LS) is a structure in the ventral forebrain that calibrates the magnitude of behavioral responses to stress-related external stimuli, including the regulation of threat avoidance. The complex connectivity between the LS and other parts of the brain, together with its largely unexplored neuronal diversity, makes it difficult to understand how defined LS circuits control specific behaviors. Here, we describe a mouse model in which a population of neurons with a common developmental origin (Nkx2.1-lineage neurons) are absent from the LS. Using a combination of circuit tracing and behavioral analyses, we found that these neurons receive inputs from the perifornical area of the anterior hypothalamus (PeFAH) and are specifically activated in stressful contexts. Mice lacking Nkx2.1-lineage LS neurons display increased exploratory behavior even under stressful conditions. Our study extends the current knowledge about how defined neuronal populations within the LS can evaluate contextual information to select appropriate behavioral responses. This is a necessary step towards understanding the crucial role that the LS plays in neuropsychiatric conditions where defensive behavior is dysregulated, such as anxiety and aggression disorders.
Figures
Similar articles
-
A newly identified mouse hypothalamic area having bidirectional neural connections with the lateral septum: the perifornical area of the anterior hypothalamus rich in chondroitin sulfate proteoglycans.Eur J Neurosci. 2015 Sep;42(6):2322-34. doi: 10.1111/ejn.13024. Epub 2015 Aug 19. Eur J Neurosci. 2015. PMID: 26205995
-
A Newly Defined Area of the Mouse Anterior Hypothalamus Involved in Septohypothalamic Circuit: Perifornical Area of the Anterior Hypothalamus, PeFAH.Acta Histochem Cytochem. 2018 Feb 27;51(1):1-8. doi: 10.1267/ahc.17030. Epub 2018 Feb 21. Acta Histochem Cytochem. 2018. PMID: 29622845 Free PMC article.
-
Lateral septum modulates cortical state to tune responsivity to threat stimuli.Cell Rep. 2022 Oct 25;41(4):111521. doi: 10.1016/j.celrep.2022.111521. Cell Rep. 2022. PMID: 36288710 Free PMC article.
-
Insights from extracellular matrix studies in the hypothalamus: structural variations of perineuronal nets and discovering a new perifornical area of the anterior hypothalamus.Anat Sci Int. 2017 Jan;92(1):18-24. doi: 10.1007/s12565-016-0375-5. Epub 2016 Oct 6. Anat Sci Int. 2017. PMID: 27714583 Review.
-
Top-down regulation of motivated behaviors via lateral septum sub-circuits.Mol Psychiatry. 2022 Aug;27(8):3119-3128. doi: 10.1038/s41380-022-01599-3. Epub 2022 May 18. Mol Psychiatry. 2022. PMID: 35581296 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources