

This is a preprint.
The H3.3 K36M oncohistone disrupts the establishment of epigenetic memory through loss of DNA methylation
- PMID: 37873347
- PMCID: PMC10592807
- DOI: 10.1101/2023.10.13.562147
The H3.3 K36M oncohistone disrupts the establishment of epigenetic memory through loss of DNA methylation
Update in
-
The H3.3K36M oncohistone disrupts the establishment of epigenetic memory through loss of DNA methylation.Mol Cell. 2024 Oct 17;84(20):3899-3915.e7. doi: 10.1016/j.molcel.2024.09.015. Epub 2024 Oct 4. Mol Cell. 2024. PMID: 39368466 Free PMC article.
Abstract
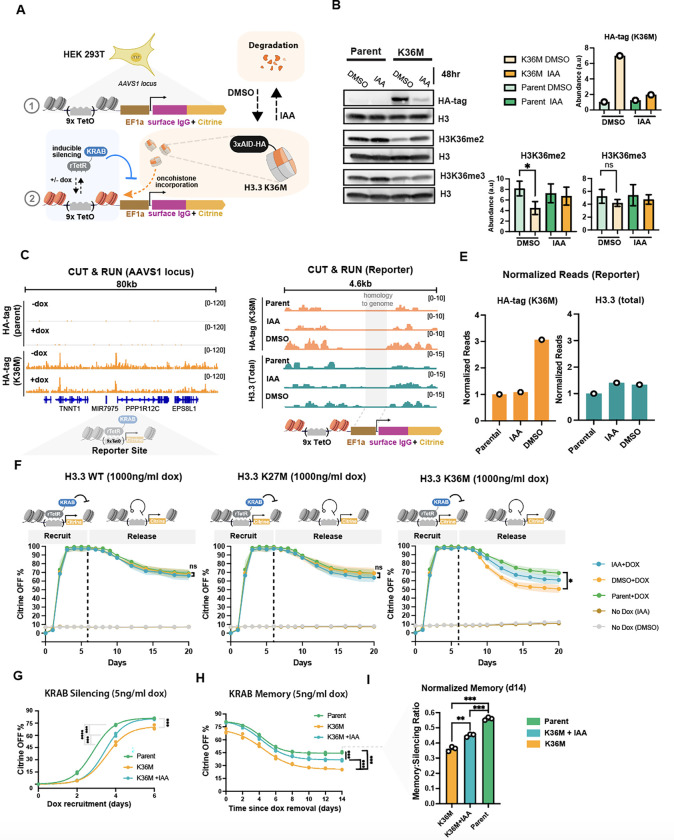
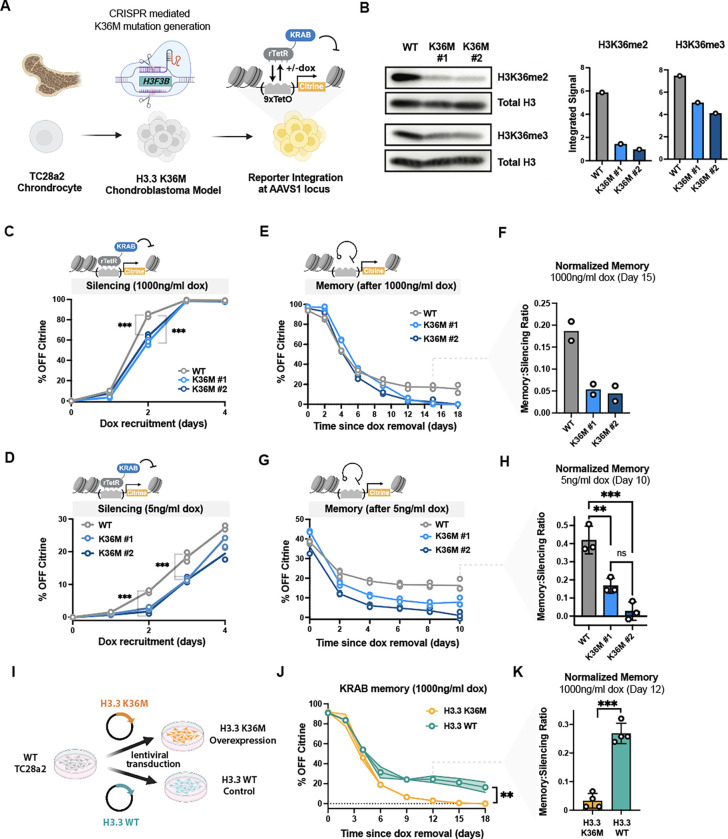
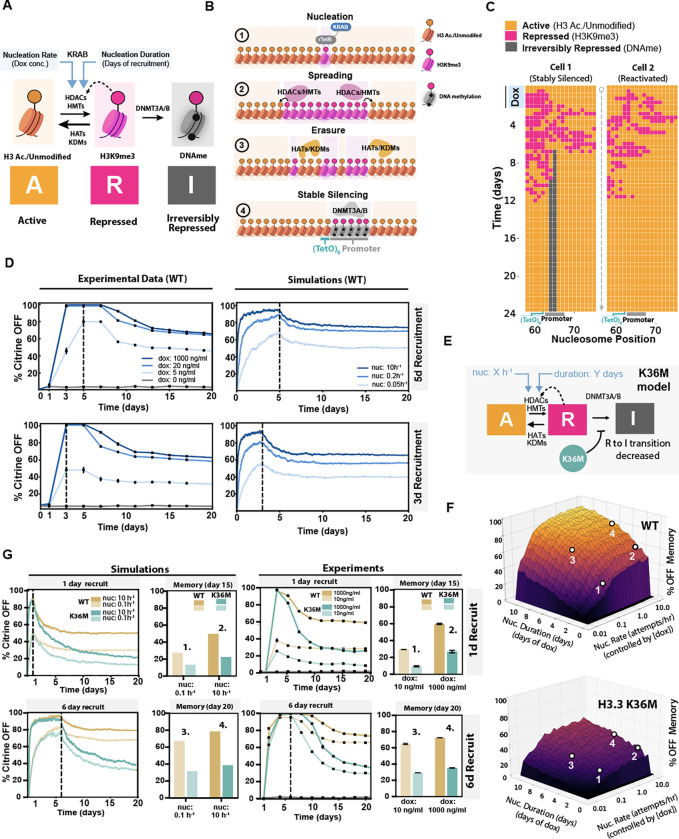
Histone H3.3 is frequently mutated in cancers, with the lysine 36 to methionine mutation (K36M) being a hallmark of chondroblastomas. While it is known that H3.3K36M changes the cellular epigenetic landscape, it remains unclear how it affects the dynamics of gene expression. Here, we use a synthetic reporter to measure the effect of H3.3K36M on silencing and epigenetic memory after recruitment of KRAB: a member of the largest class of human repressors, commonly used in synthetic biology, and associated with H3K9me3. We find that H3.3K36M, which decreases H3K36 methylation, leads to a decrease in epigenetic memory and promoter methylation weeks after KRAB release. We propose a new model for establishment and maintenance of epigenetic memory, where H3K36 methylation is necessary to convert H3K9me3 domains into DNA methylation for stable epigenetic memory. Our quantitative model can inform oncogenic mechanisms and guide development of epigenetic editing tools.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials