

The anaphase-promoting complex controls a ubiquitination-phosphoprotein axis in chromatin during neurodevelopment
- PMID: 37875116
- PMCID: PMC10872926
- DOI: 10.1016/j.devcel.2023.10.002
The anaphase-promoting complex controls a ubiquitination-phosphoprotein axis in chromatin during neurodevelopment
Abstract
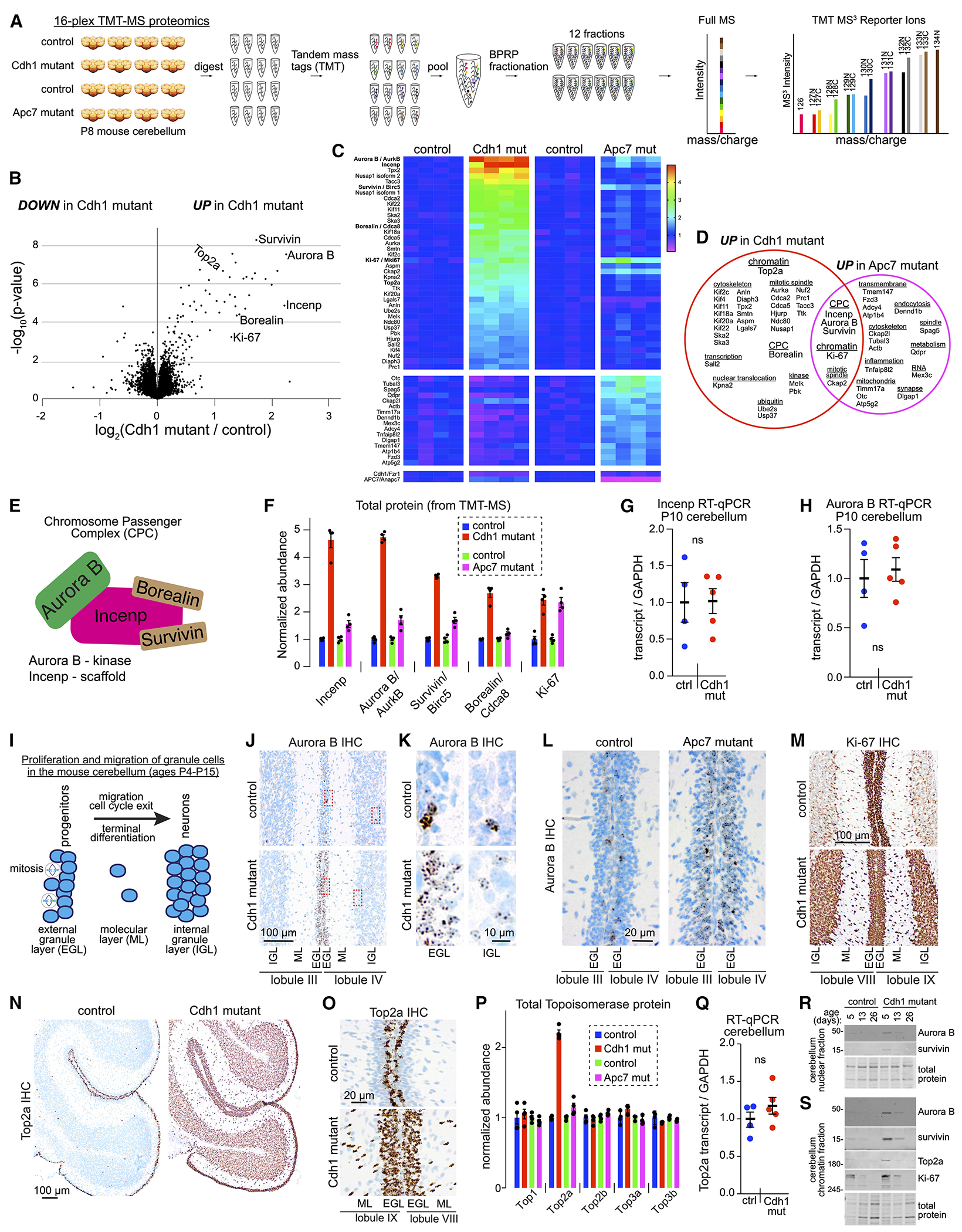
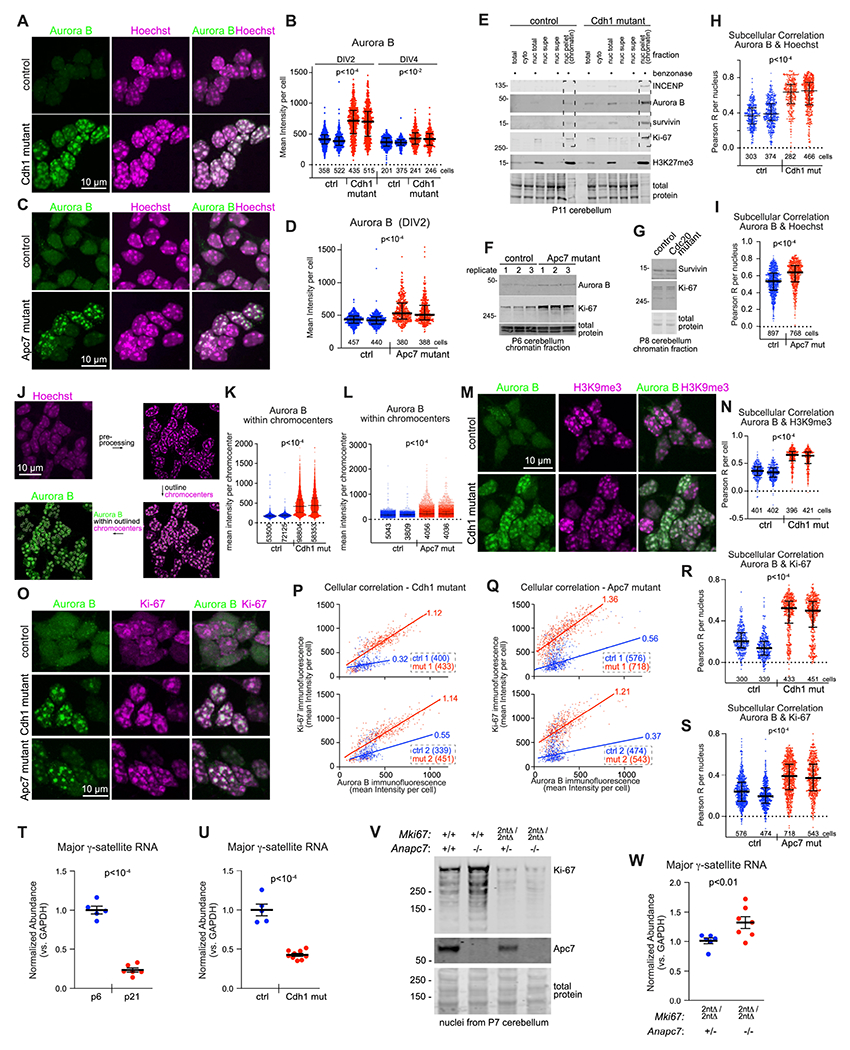
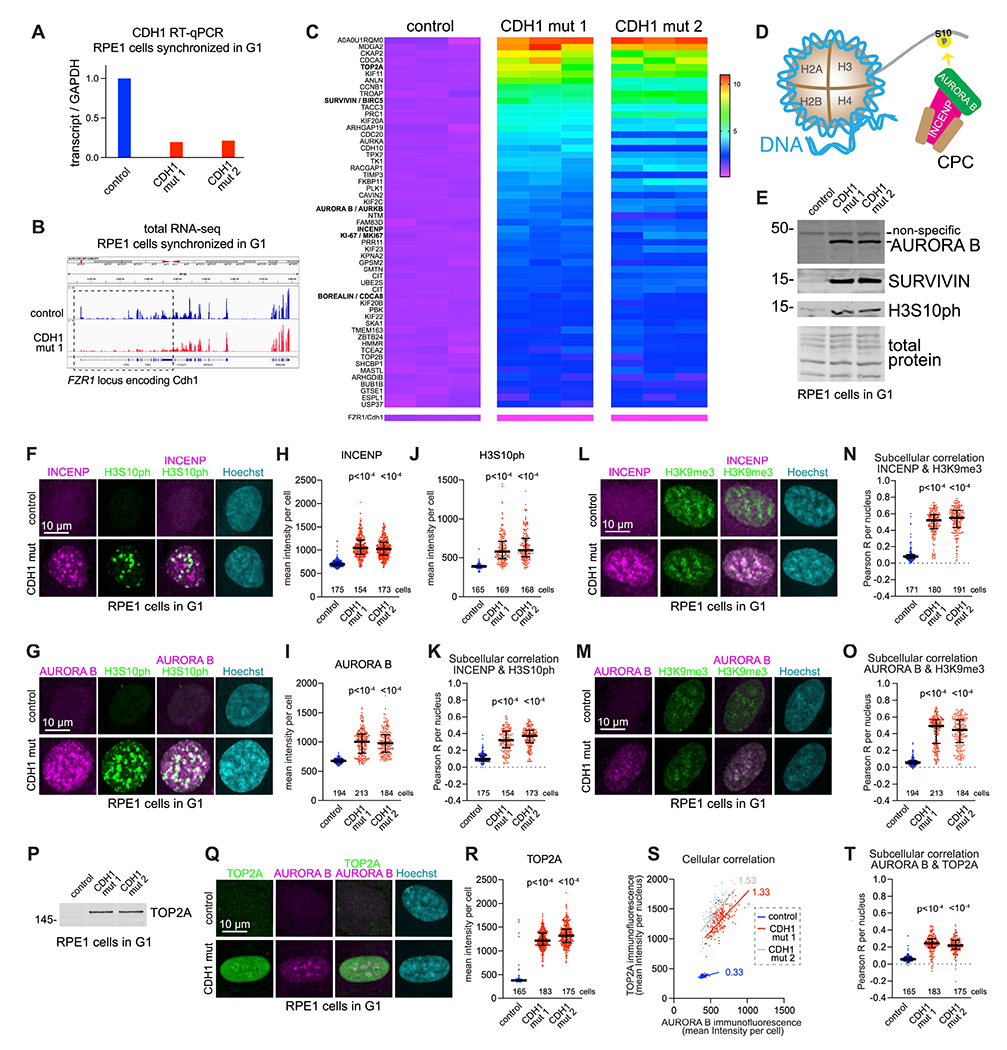
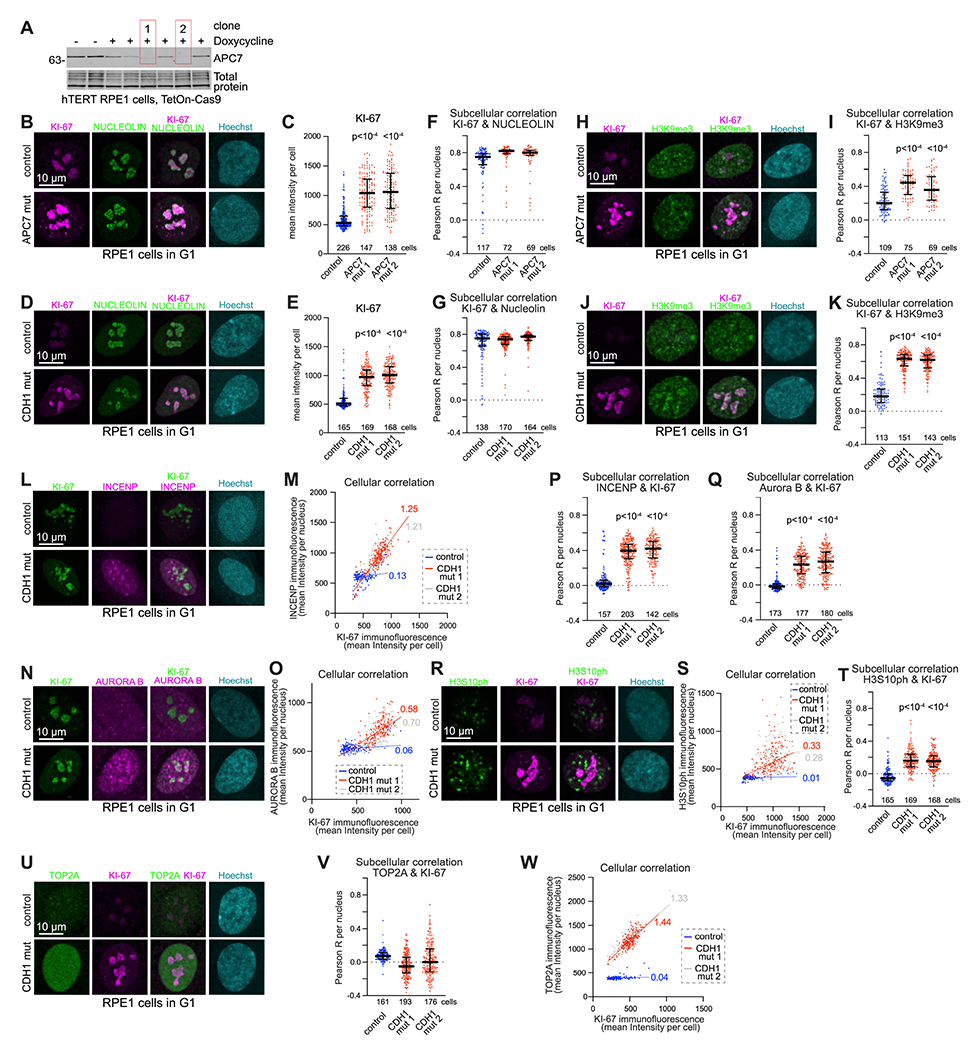
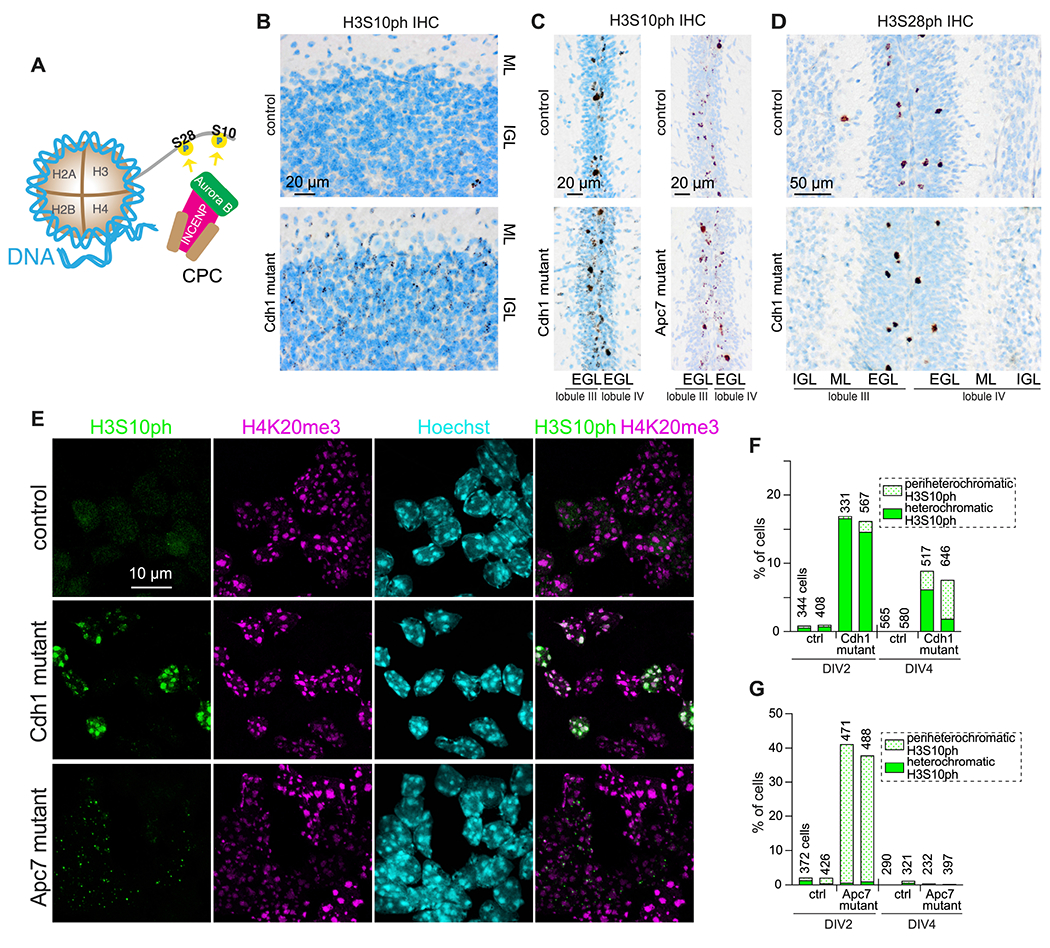
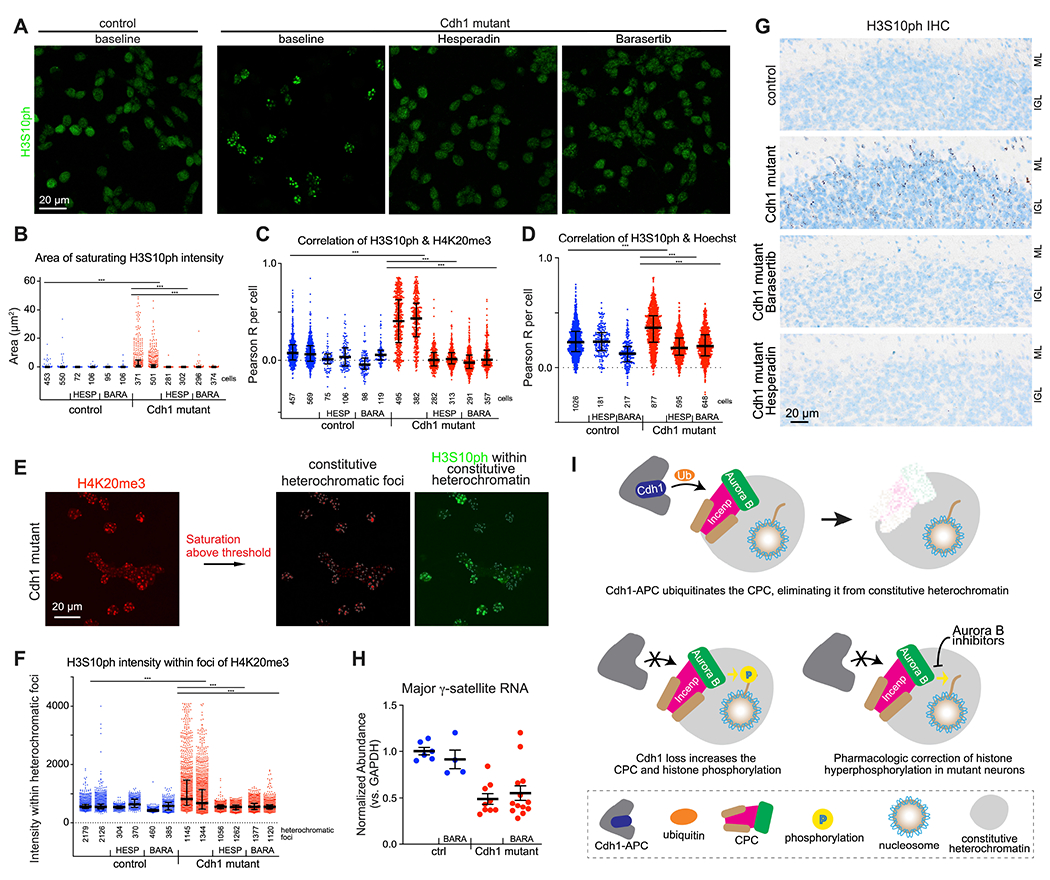
Mutations in the degradative ubiquitin ligase anaphase-promoting complex (APC) alter neurodevelopment by impairing proteasomal protein clearance, but our understanding of their molecular and cellular pathogenesis remains limited. Here, we employ the proteomic-based discovery of APC substrates in APC mutant mouse brain and human cell lines and identify the chromosome-passenger complex (CPC), topoisomerase 2a (Top2a), and Ki-67 as major chromatin factors targeted by the APC during neuronal differentiation. These substrates accumulate in phosphorylated form, suggesting that they fail to be eliminated after mitosis during terminal differentiation. The accumulation of the CPC kinase Aurora B within constitutive heterochromatin and hyperphosphorylation of its target histone 3 are corrected in the mutant brain by pharmacologic Aurora B inhibition. Surprisingly, the reduction of Ki-67, but not H3S10ph, rescued the function of constitutive heterochromatin in APC mutant neurons. These results expand our understanding of how ubiquitin signaling regulates chromatin during neurodevelopment and identify potential therapeutic targets in APC-related disorders.
Keywords: H3S10ph; Ki-67; anaphase-promoting complex; chromatin; chromosome-passenger complex; heterochromatin; neurodevelopment; proteomics/phosphoproteomics; topoisomerase; ubiquitin ligase.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

References
-
- Dikic I. (2017). Proteasomal and autophagic degradation systems. Annu. Rev. Biochem 86, 193–224. - PubMed
-
- Pohl C & Dikic I (2019). Cellular quality control by the ubiquitin-proteasome system and autophagy. Science. 366, 818–822. - PubMed
-
- Deshaies RJ & Joazeiro CAP (2009). RING domain E3 ubiquitin ligases. Annu. Rev. Biochem 78, 399–434. - PubMed
-
- Buiting K, Williams C & Horsthemke B (2016). Angelman syndrome-insights into a rare neurogenetic disorder. Nat. Rev. Neurol 12, 584–593. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
