

Reducing cell intrinsic immunity to mRNA vaccine alters adaptive immune responses in mice
- PMID: 37876532
- PMCID: PMC10591005
- DOI: 10.1016/j.omtn.2023.102045
Reducing cell intrinsic immunity to mRNA vaccine alters adaptive immune responses in mice
Abstract
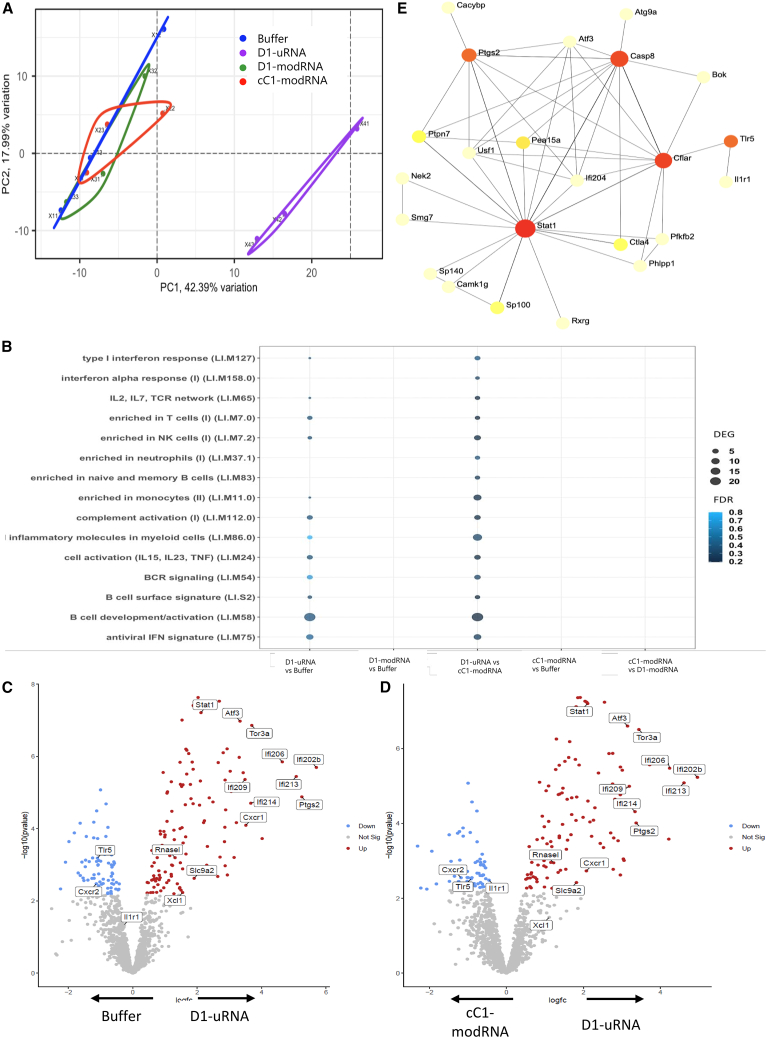
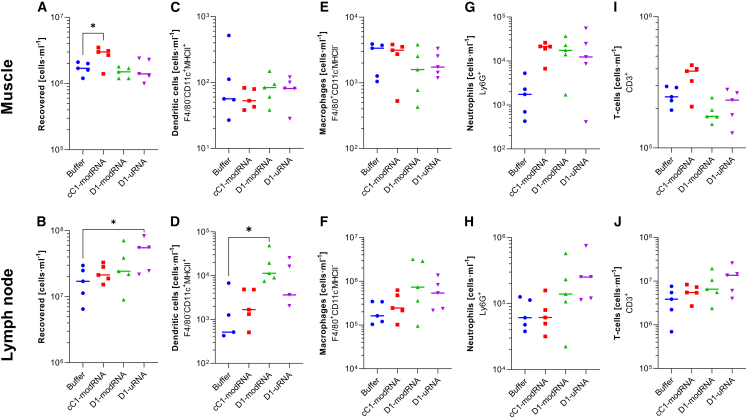
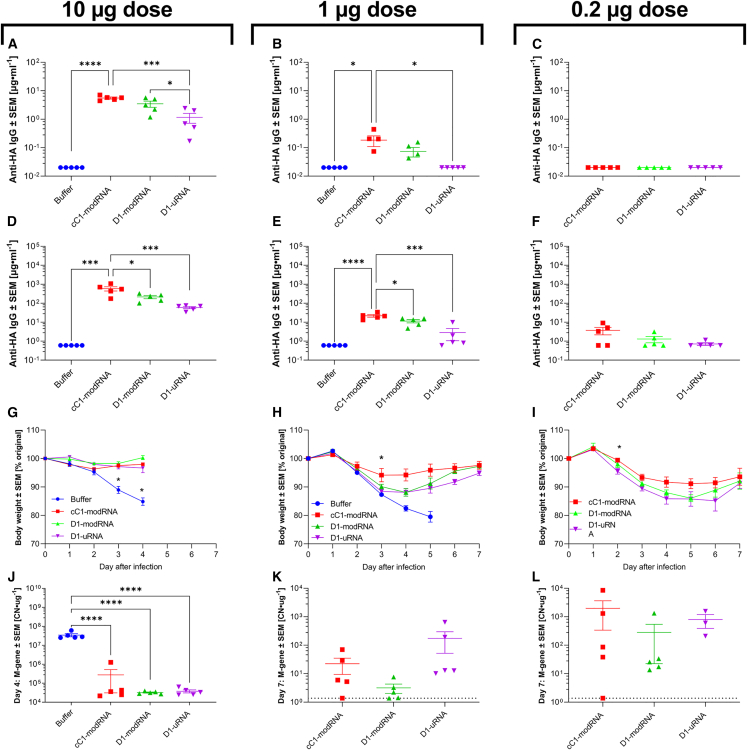
The response to mRNA vaccines needs to be sufficient for immune cell activation and recruitment, but moderate enough to ensure efficacious antigen expression. The choice of the cap structure and use of N1-methylpseudouridine (m1Ψ) instead of uridine, which have been shown to reduce RNA sensing by the cellular innate immune system, has led to improved efficacy of mRNA vaccine platforms. Understanding how RNA modifications influence the cell intrinsic immune response may help in the development of more effective mRNA vaccines. In the current study, we compared mRNA vaccines in mice against influenza virus using three different mRNA formats: uridine-containing mRNA (D1-uRNA), m1Ψ-modified mRNA (D1-modRNA), and D1-modRNA with a cap1 structure (cC1-modRNA). D1-uRNA vaccine induced a significantly different gene expression profile to the modified mRNA vaccines, with an up-regulation of Stat1 and RnaseL, and increased systemic inflammation. This result correlated with significantly reduced antigen-specific antibody responses and reduced protection against influenza virus infection compared with D1-modRNA and cC1-modRNA. Incorporation of m1Ψ alone without cap1 improved antibodies, but both modifications were required for the optimum response. Therefore, the incorporation of m1Ψ and cap1 alters protective immunity from mRNA vaccines by altering the innate immune response to the vaccine material.
Keywords: MT: Bioinformatics; RNA vaccine; cell intrinsic; inflammation; influenza; innate; sensing.
© 2023 The Authors.
Conflict of interest statement
E.J.J., S.K., J.M., L.A.S., A.B.V., K.K., U.S., and S.E. are employees at BioNTech SE (Mainz, Germany). U.S. is cofounder and management board member of BioNTech SE (Mainz, Germany). A.B.V., K.K., U.S., and S.E. are inventors on patents and patent applications related to RNA technology. A.B.V., K.K., U.S., and S.E. hold securities from BioNTech SE.
Figures





References
-
- Sahin U., Muik A., Vogler I., Derhovanessian E., Kranz L.M., Vormehr M., Quandt J., Bidmon N., Ulges A., Baum A., et al. BNT162b2 vaccine induces neutralizing antibodies and poly-specific T cells in humans. Nature. 2021;595:572–577. - PubMed
-
- Karikó K., Buckstein M., Ni H., Weissman D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity. 2005;23:165–175. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
