

Pre- and postsynaptic N-methyl-D-aspartate receptors are required for sequential printing of fear memory engrams
- PMID: 37876798
- PMCID: PMC10590821
- DOI: 10.1016/j.isci.2023.108050
Pre- and postsynaptic N-methyl-D-aspartate receptors are required for sequential printing of fear memory engrams
Abstract
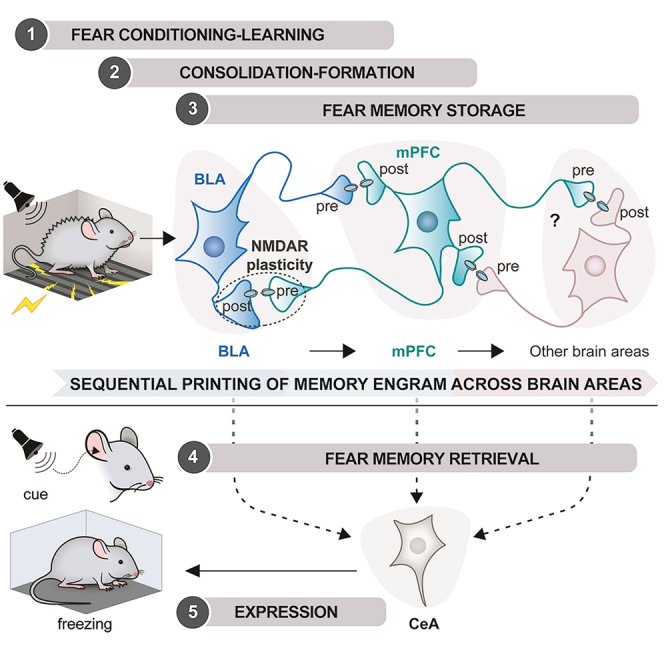
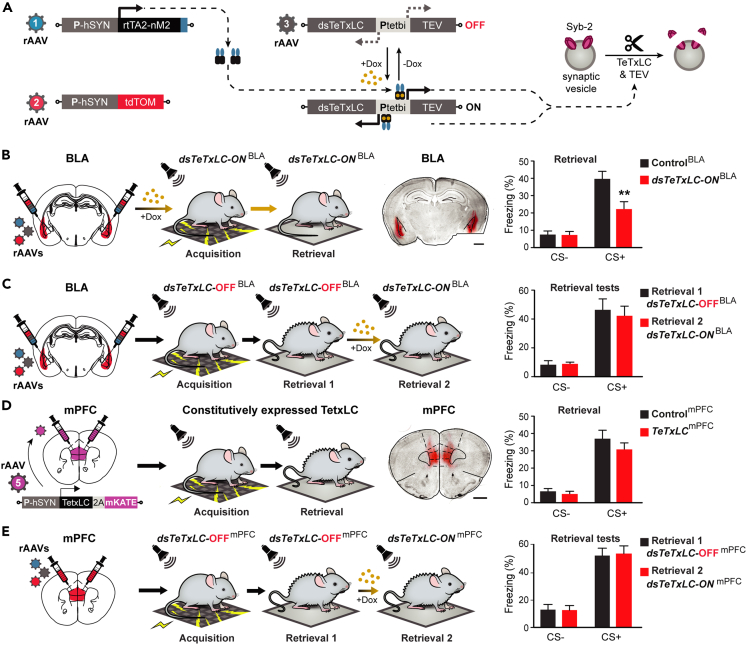
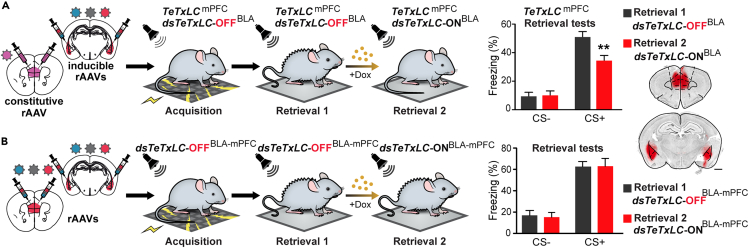
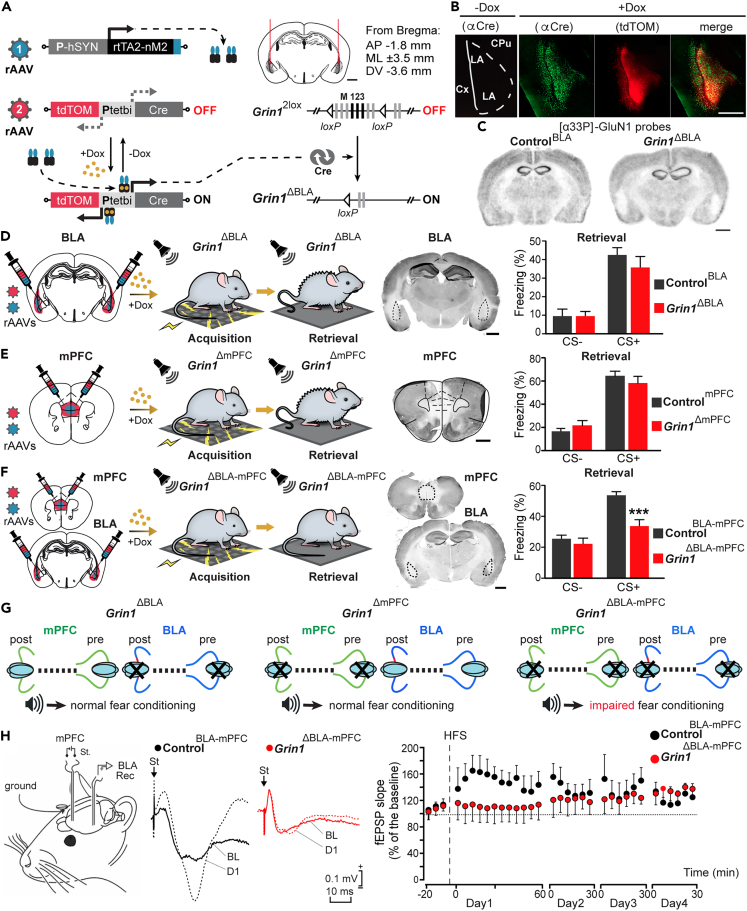
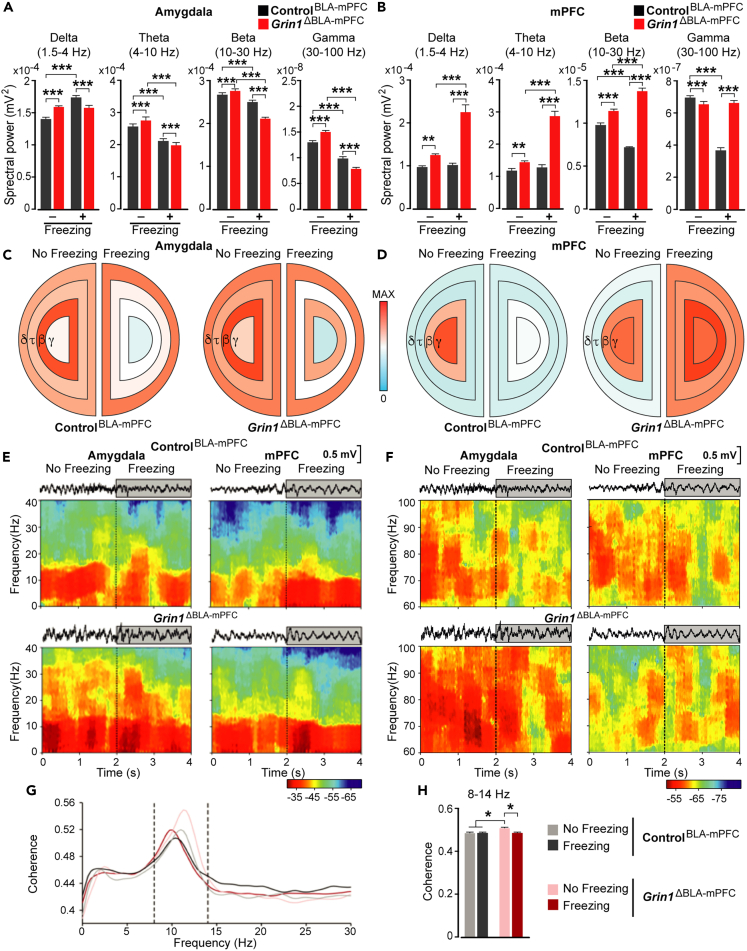
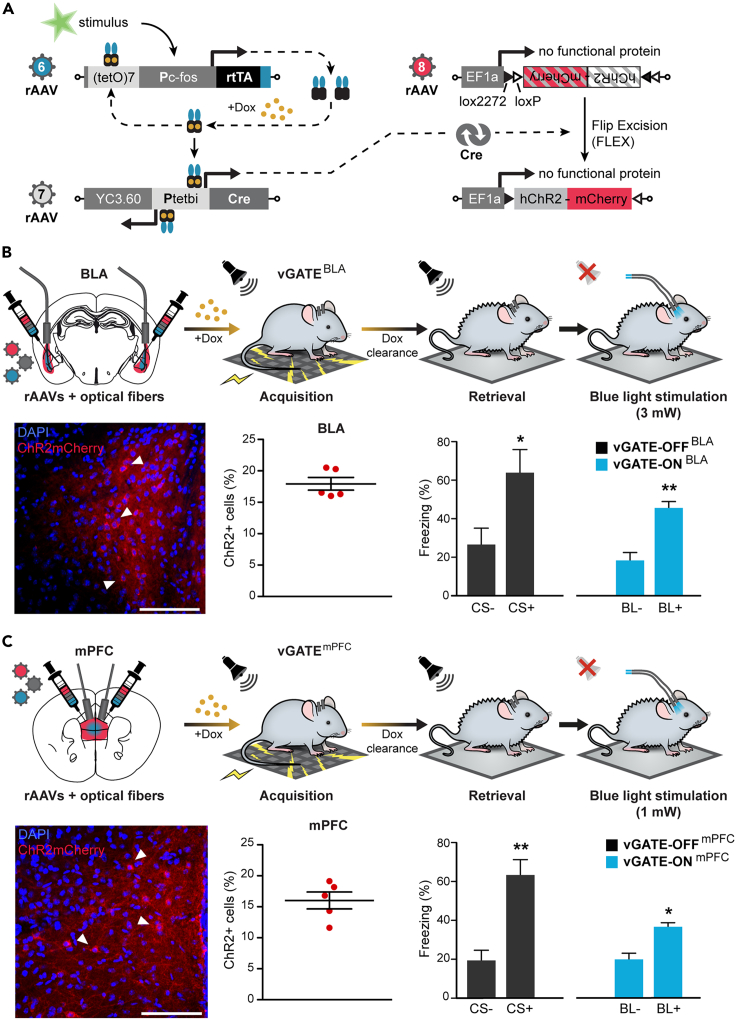
The organization of fear memory involves the participation of multiple brain regions. However, it is largely unknown how fear memory is formed, which circuit pathways are used for "printing" memory engrams across brain regions, and the role of identified brain circuits in memory retrieval. With advanced genetic methods, we combinatorially blocked presynaptic output and manipulated N-methyl-D-aspartate receptor (NMDAR) in the basolateral amygdala (BLA) and medial prefrontal cortex (mPFC) before and after cued fear conditioning. Further, we tagged fear-activated neurons during associative learning for optogenetic memory recall. We found that presynaptic mPFC and postsynaptic BLA NMDARs are required for fear memory formation, but not expression. Our results provide strong evidence that NMDAR-dependent synaptic plasticity drives multi-trace systems consolidation for the sequential printing of fear memory engrams from BLA to mPFC and, subsequently, to the other regions, for flexible memory retrieval.
Keywords: Behavioral neuroscience; Cellular neuroscience.
© 2023.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Tulving E. Multiple memory systems and consciousness. Hum. Neurobiol. 1987;6:67–80. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
