

Synaptic BMAL1 phosphorylation controls circadian hippocampal plasticity
- PMID: 37878694
- PMCID: PMC10599629
- DOI: 10.1126/sciadv.adj1010
Synaptic BMAL1 phosphorylation controls circadian hippocampal plasticity
Abstract
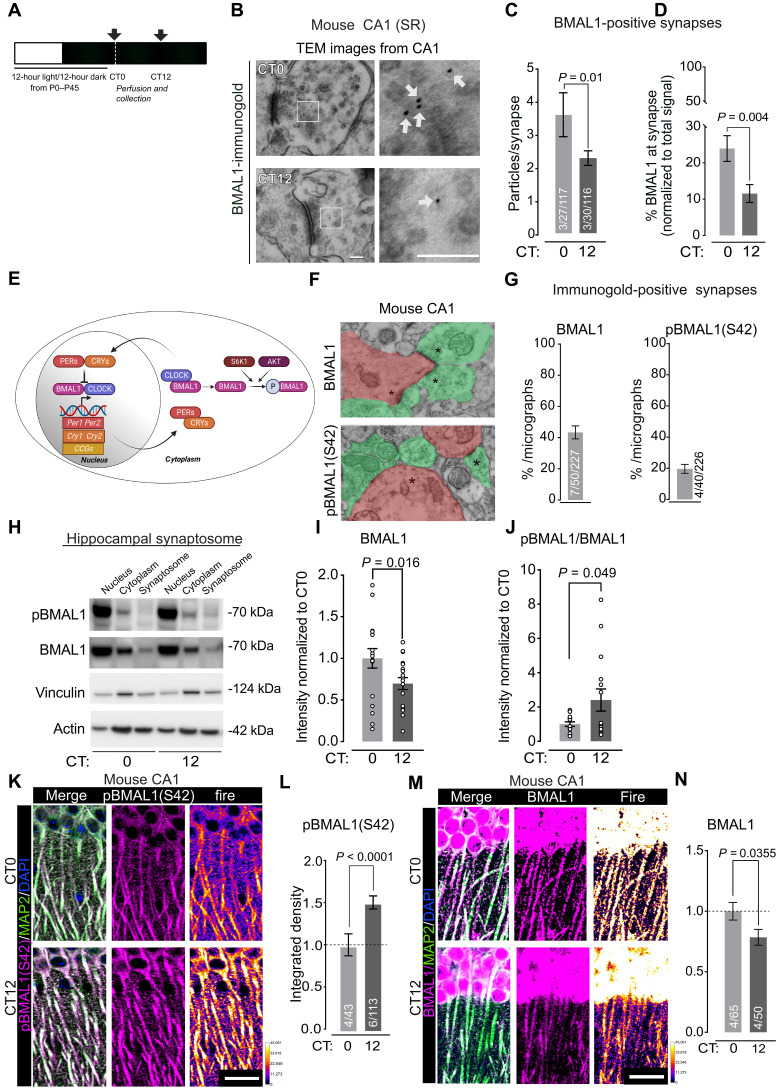
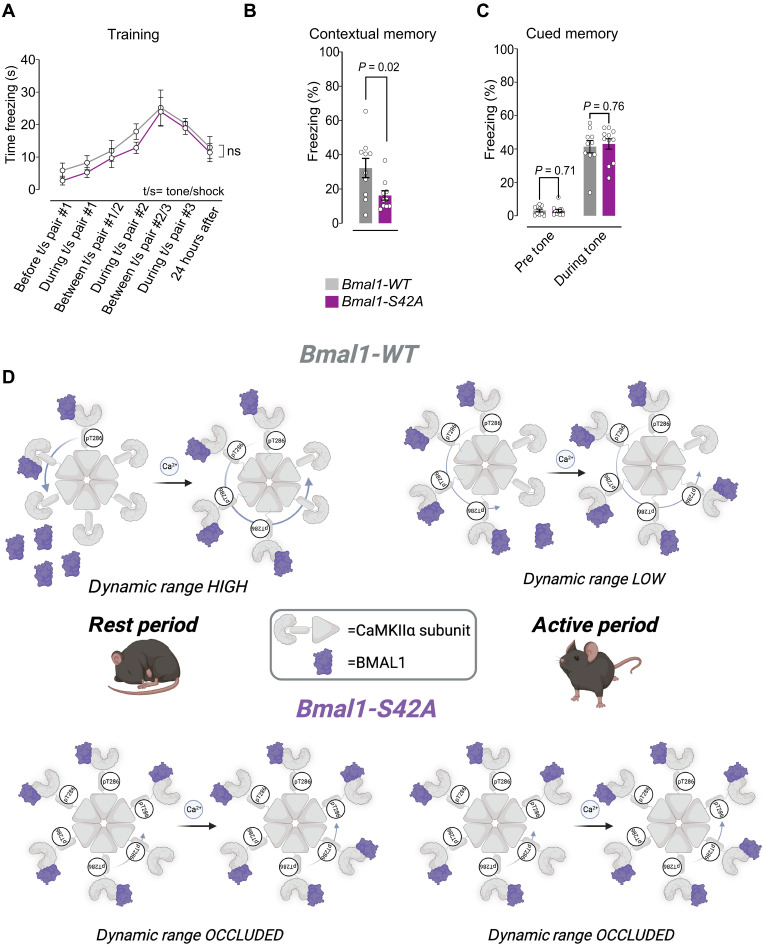
The time of day strongly influences adaptive behaviors like long-term memory, but the correlating synaptic and molecular mechanisms remain unclear. The circadian clock comprises a canonical transcription-translation feedback loop (TTFL) strictly dependent on the BMAL1 transcription factor. We report that BMAL1 rhythmically localizes to hippocampal synapses in a manner dependent on its phosphorylation at Ser42 [pBMAL1(S42)]. pBMAL1(S42) regulates the autophosphorylation of synaptic CaMKIIα and circadian rhythms of CaMKIIα-dependent molecular interactions and LTP but not global rest/activity behavior. Therefore, our results suggest a model in which repurposing of the clock protein BMAL1 to synapses locally gates the circadian timing of plasticity.
Figures





References
-
- C. Gizowski, C. Zaelzer, C. W. Bourque, Clock-driven vasopressin neurotransmission mediates anticipatory thirst prior to sleep. Nature 537, 685–688 (2016). - PubMed
-
- P. Valnegri, M. Khelfaoui, O. Dorseuil, S. Bassani, C. Lagneaux, A. Gianfelice, R. Benfante, J. Chelly, P. Billuart, C. Sala, M. Passafaro, A circadian clock in hippocampus is regulated by interaction between oligophrenin-1 and Rev-erbα. Nat. Neurosci. 14, 1293–1301 (2011). - PubMed
MeSH terms
Substances
Grants and funding
- T32 GM007753/GM/NIGMS NIH HHS/United States
- R01 HL151368/HL/NHLBI NIH HHS/United States
- R01 MH111647/MH/NIMH NIH HHS/United States
- RF1 NS126547/NS/NINDS NIH HHS/United States
- R21 MH120438/MH/NIMH NIH HHS/United States
- R43 NS119081/NS/NINDS NIH HHS/United States
- U44 NS121559/NS/NINDS NIH HHS/United States
- R01 MH120069/MH/NIMH NIH HHS/United States
- T32 HL139438/HL/NHLBI NIH HHS/United States
- R21 NS123459/NS/NINDS NIH HHS/United States
- P50 HD105351/HD/NICHD NIH HHS/United States
- F30 MH132277/MH/NIMH NIH HHS/United States
- T32 GM144273/GM/NIGMS NIH HHS/United States
- R21 NS121858/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
