

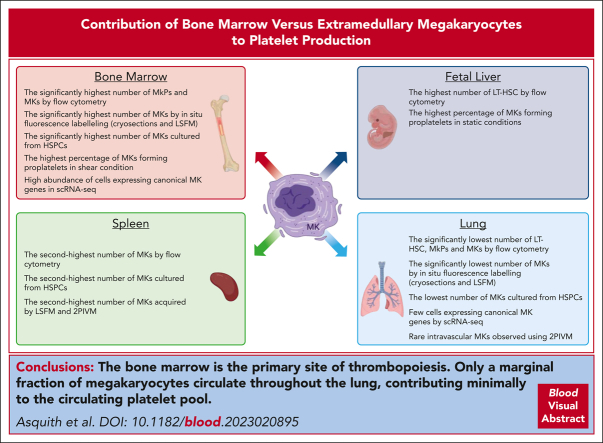
The bone marrow is the primary site of thrombopoiesis
- PMID: 37879046
- PMCID: PMC10808241
- DOI: 10.1182/blood.2023020895
The bone marrow is the primary site of thrombopoiesis
Abstract
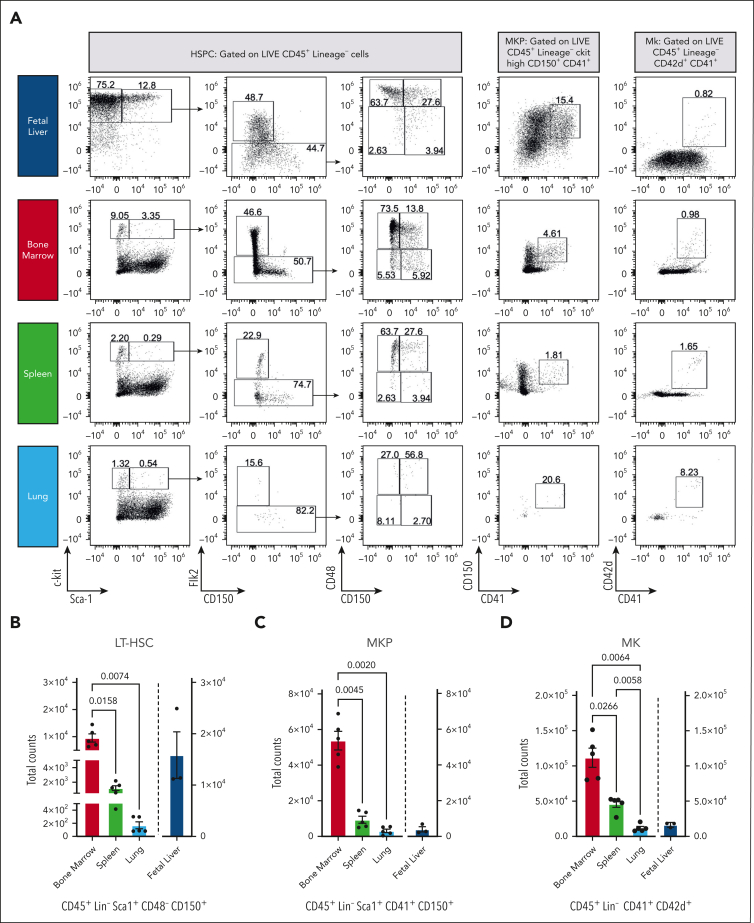
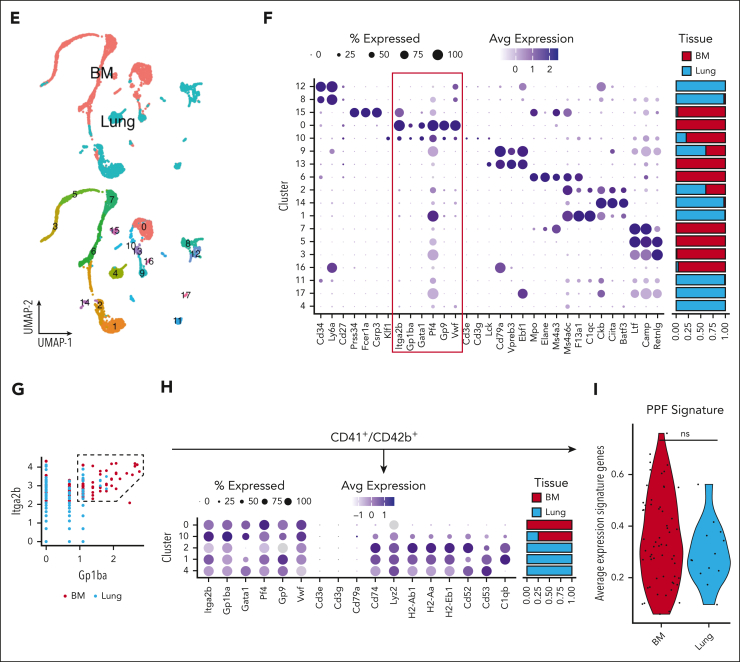
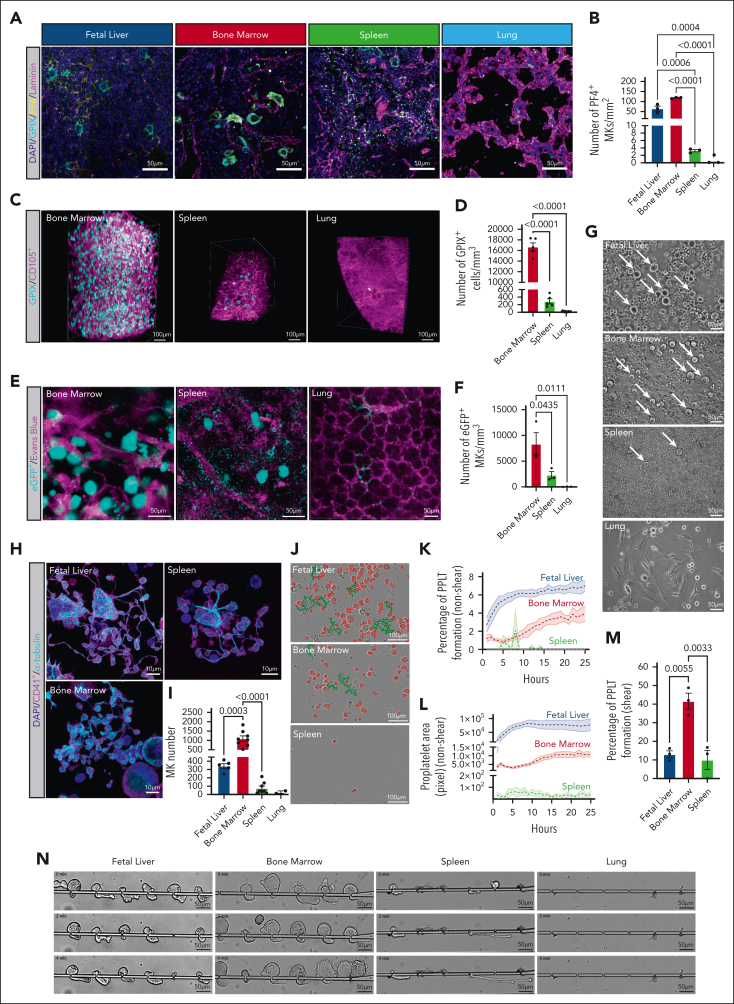
Megakaryocytes (MKs) generate thousands of platelets over their lifespan. The roles of platelets in infection and inflammation has guided an interest to the study of extramedullary thrombopoiesis and therefore MKs have been increasingly reported within the spleen and lung. However, the relative abundance of MKs in these organs compared to the bone marrow and the scale of their contribution to the platelet pool in a steady state remain controversial. We investigated the relative abundance of MKs in the adult murine bone marrow, spleen, and lung using whole-mount light-sheet and quantitative histological imaging, flow cytometry, intravital imaging, and an assessment of single-cell RNA sequencing (scRNA-seq) repositories. Flow cytometry revealed significantly higher numbers of hematopoietic stem and progenitor cells and MKs in the murine bone marrow than in spleens or perfused lungs. Two-photon intravital and light-sheet microscopy, as well as quantitative histological imaging, confirmed these findings. Moreover, ex vivo cultured MKs from the bone marrow subjected to static or microfluidic platelet production assays had a higher capacity for proplatelet formation than MKs from other organs. Analysis of previously published murine and human scRNA-seq data sets revealed that only a marginal fraction of MK-like cells can be found within the lung and most likely only marginally contribute to platelet production in the steady state.
© 2024 American Society of Hematology. Published by Elsevier Inc. All rights are reserved, including those for text and data mining, AI training, and similar technologies.
Conflict of interest statement
Conflict-of-interest disclosure: J.E.I. has financial interest in and is a founder of StellularBio, a biotechnology company focused on making donor-independent platelet-like cells at scale; these interests are managed by Boston Children’s Hospital. The remaining authors declare no competing financial interests.
Figures
Comment in
-
Megakaryocytes in the lung: guests or ghosts?Blood. 2024 Jan 18;143(3):192-193. doi: 10.1182/blood.2023022897. Blood. 2024. PMID: 38236613 No abstract available.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
