

Structures of a sperm-specific solute carrier gated by voltage and cAMP
- PMID: 37880361
- PMCID: PMC10620091
- DOI: 10.1038/s41586-023-06629-w
Structures of a sperm-specific solute carrier gated by voltage and cAMP
Abstract
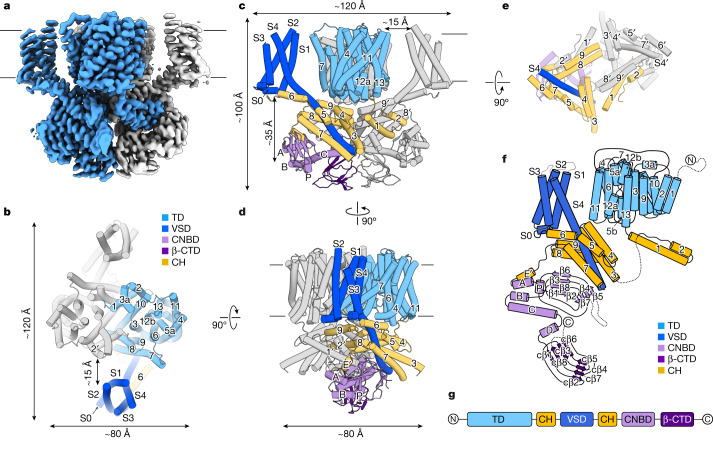
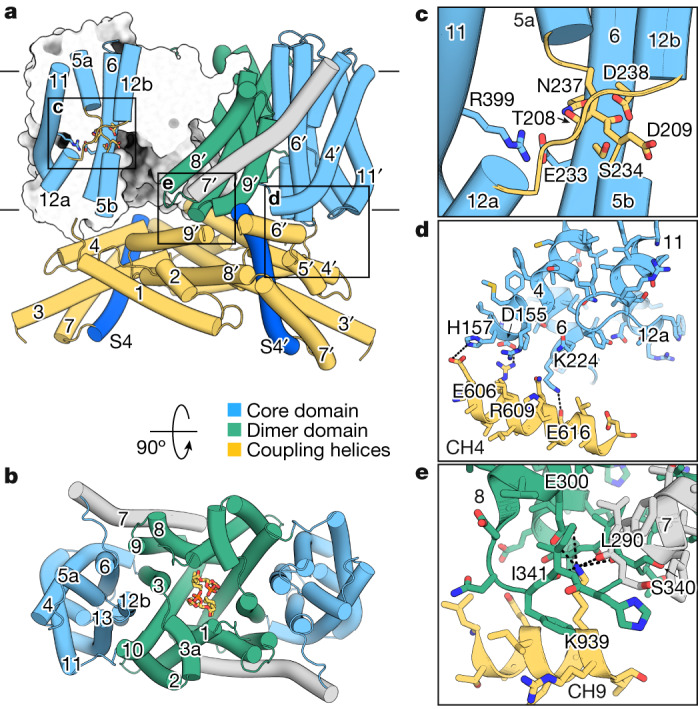
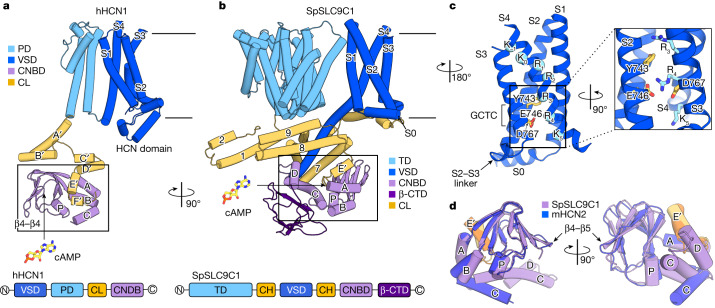
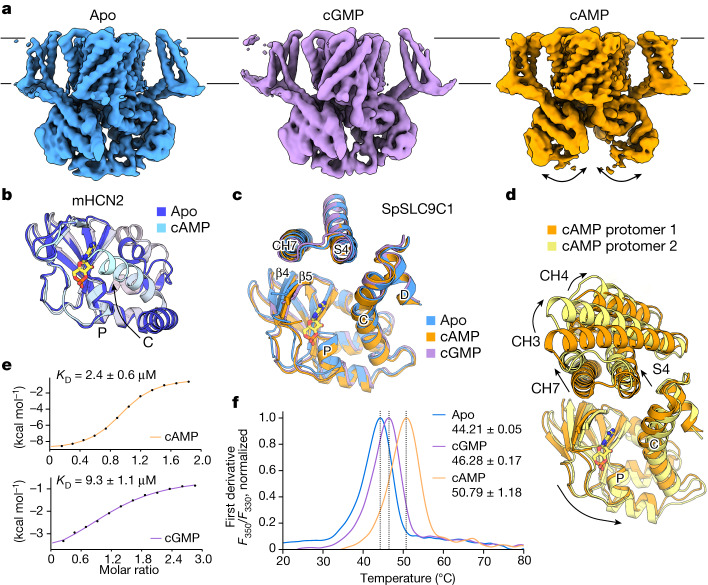
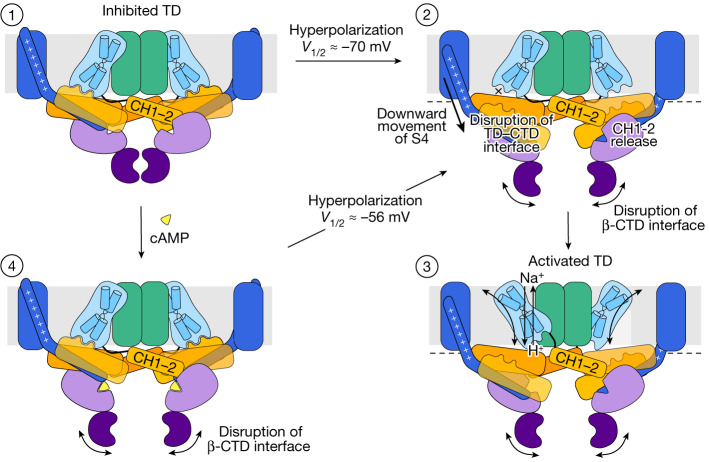
The newly characterized sperm-specific Na+/H+ exchanger stands out by its unique tripartite domain composition1,2. It unites a classical solute carrier unit with regulatory domains usually found in ion channels, namely, a voltage-sensing domain and a cyclic-nucleotide binding domain1,3, which makes it a mechanistic chimera and a secondary-active transporter activated strictly by membrane voltage. Our structures of the sea urchin SpSLC9C1 in the absence and presence of ligands reveal the overall domain arrangement and new structural coupling elements. They allow us to propose a gating model, where movements in the voltage sensor indirectly cause the release of the exchanging unit from a locked state through long-distance allosteric effects transmitted by the newly characterized coupling helices. We further propose that modulation by its ligand cyclic AMP occurs by means of disruption of the cytosolic dimer interface, which lowers the energy barrier for S4 movements in the voltage-sensing domain. As SLC9C1 members have been shown to be essential for male fertility, including in mammals2,4,5, our structure represents a potential new platform for the development of new on-demand contraceptives.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures








Comment in
-
An ion transporter in sperm that has features of a channel.Nature. 2023 Nov;623(7985):38-40. doi: 10.1038/d41586-023-03154-8. Nature. 2023. PMID: 37880527 No abstract available.
References
-
- Cavarocchi, E. et al. The sodium/proton exchanger SLC9C1 (sNHE) is essential for human sperm motility and fertility. Clin. Genet.10.1111/cge.13927 (2021). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
