

Dopaminergic systems create reward seeking despite adverse consequences
- PMID: 37880370
- PMCID: PMC10632144
- DOI: 10.1038/s41586-023-06671-8
Dopaminergic systems create reward seeking despite adverse consequences
Erratum in
-
Publisher Correction: Dopaminergic systems create reward seeking despite adverse consequences.Nature. 2023 Dec;624(7991):E5. doi: 10.1038/s41586-023-06876-x. Nature. 2023. PMID: 38012376 Free PMC article. No abstract available.
Abstract
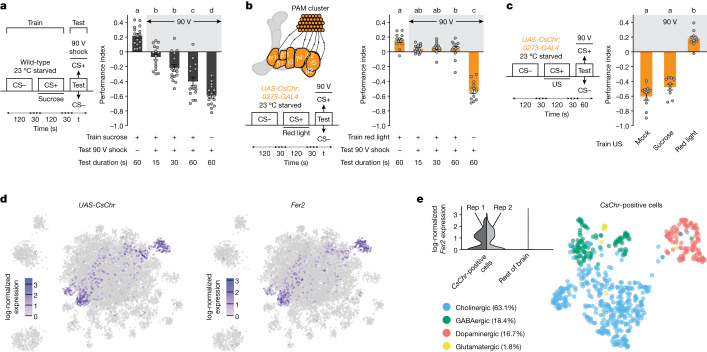
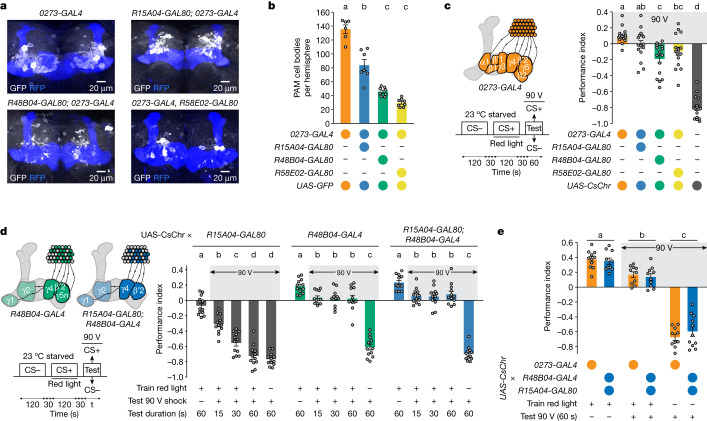
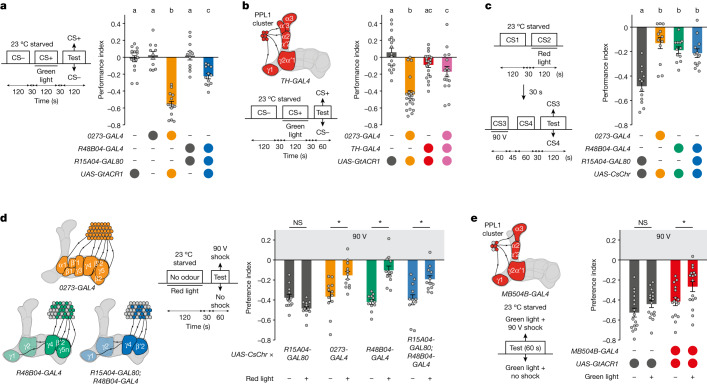
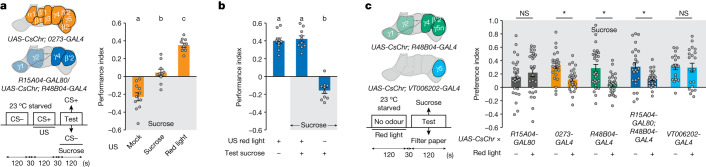
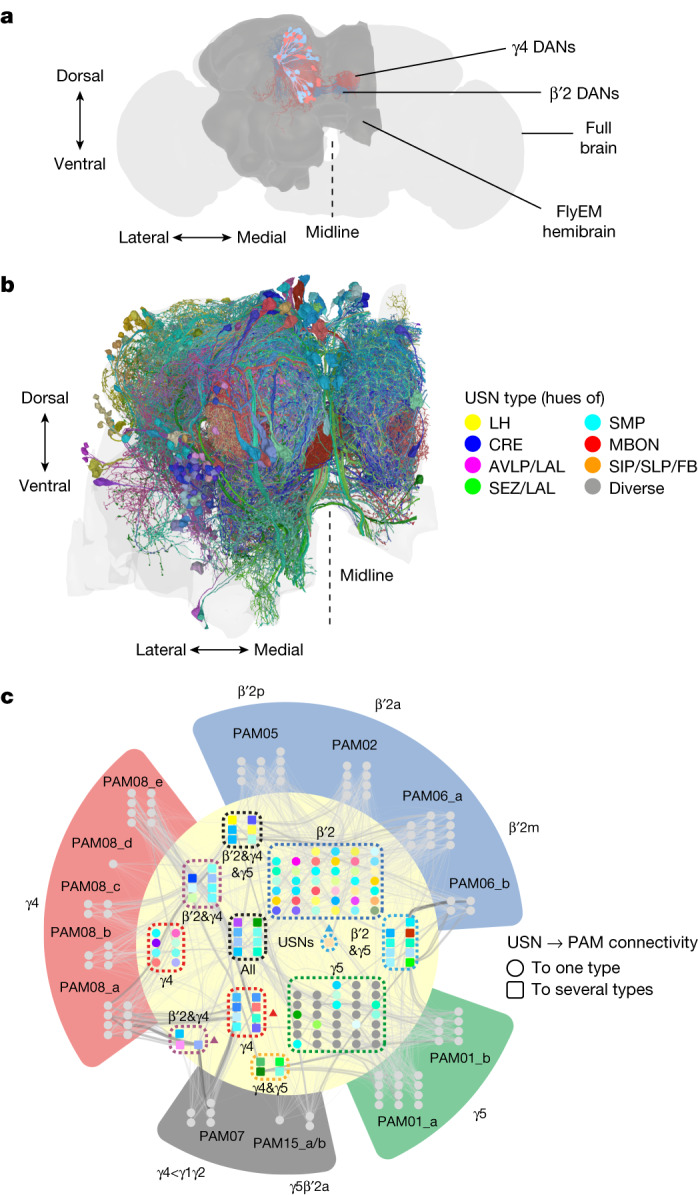
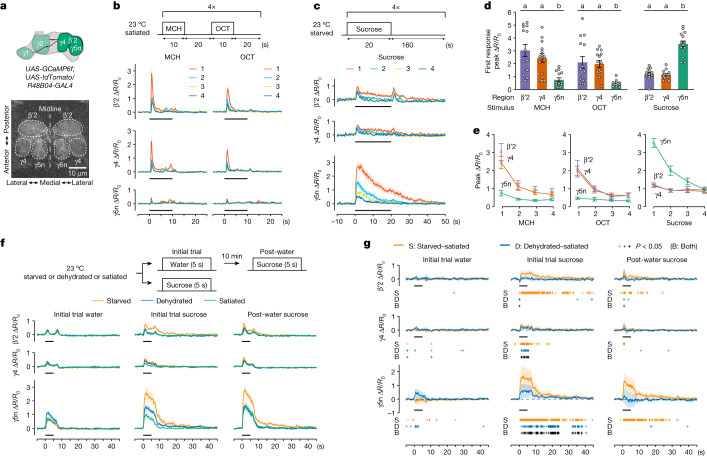
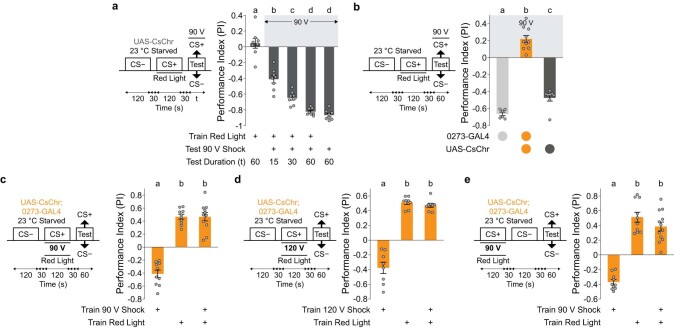
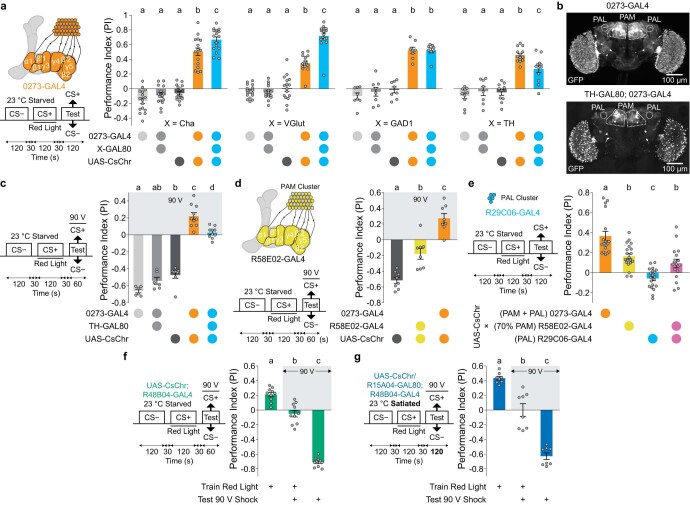
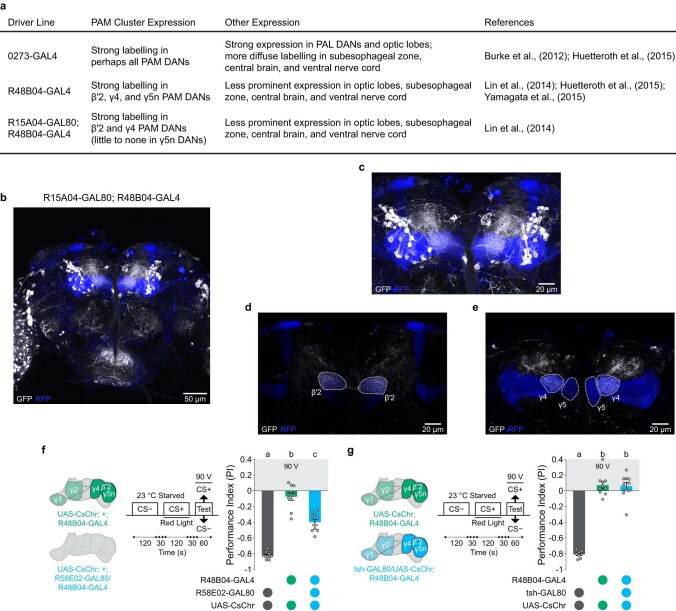
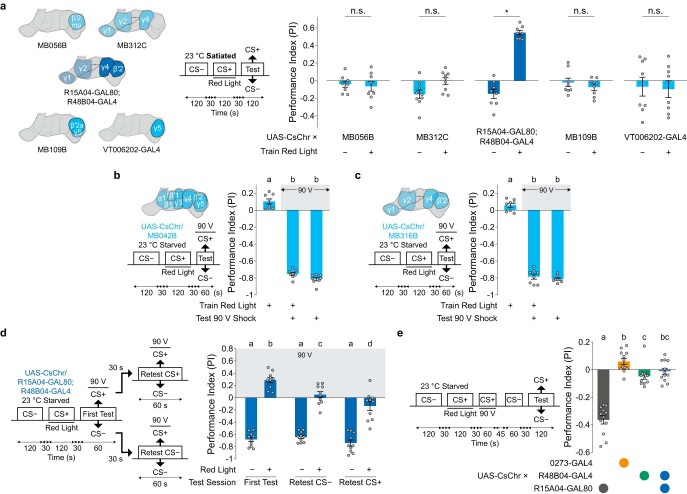
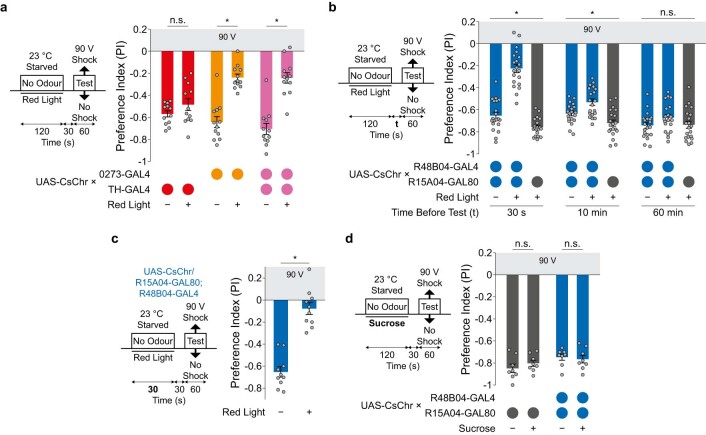
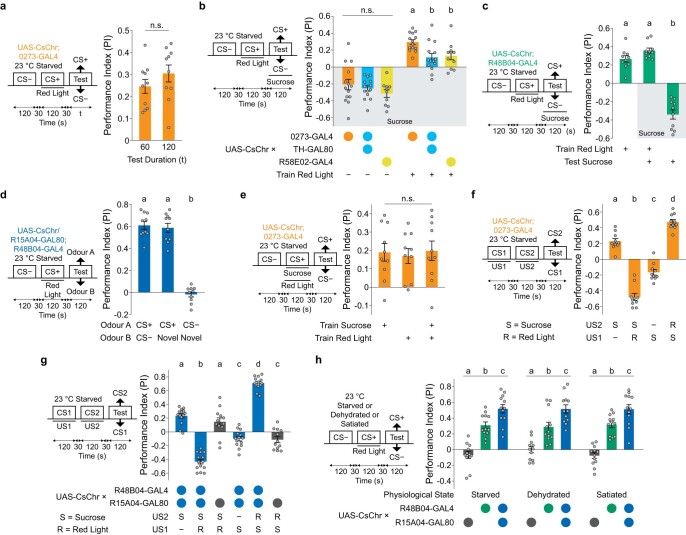
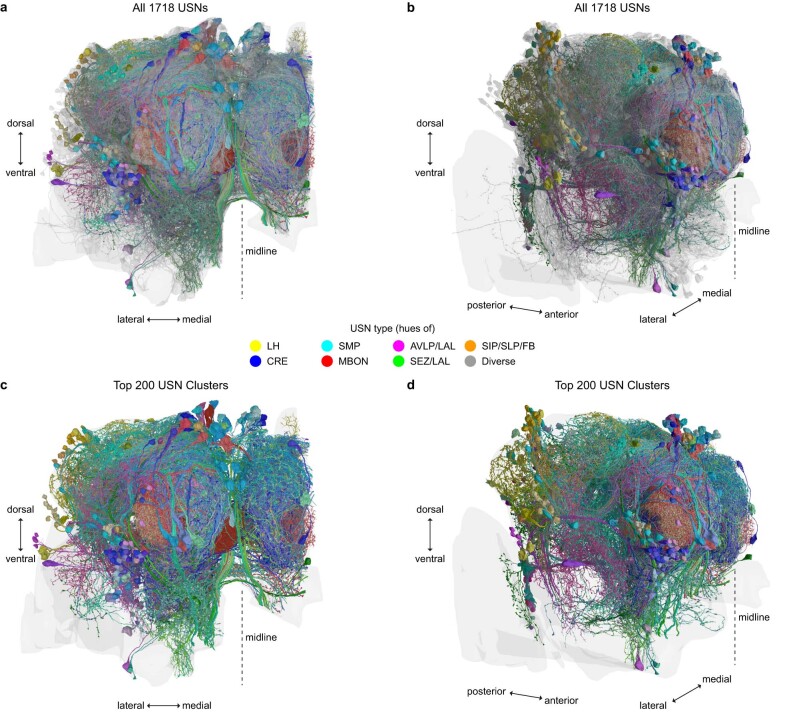
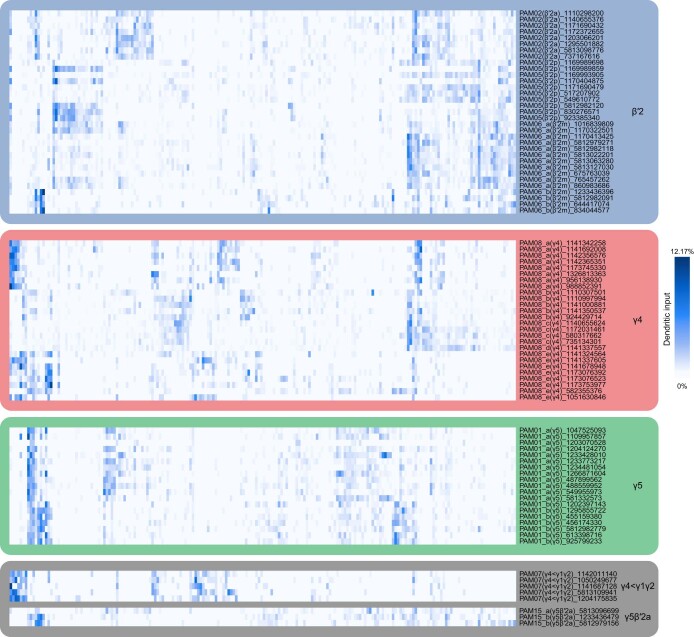
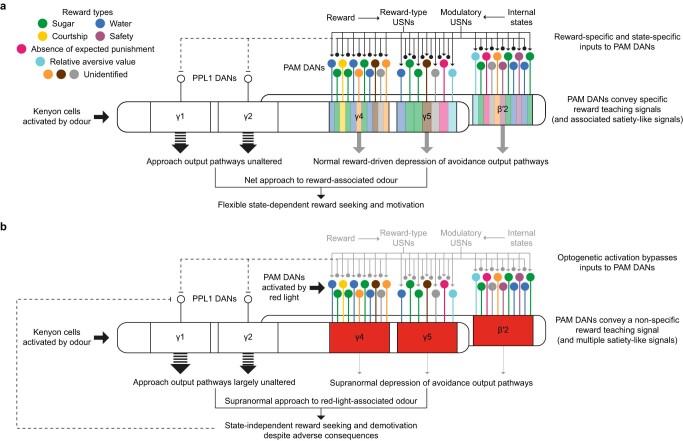
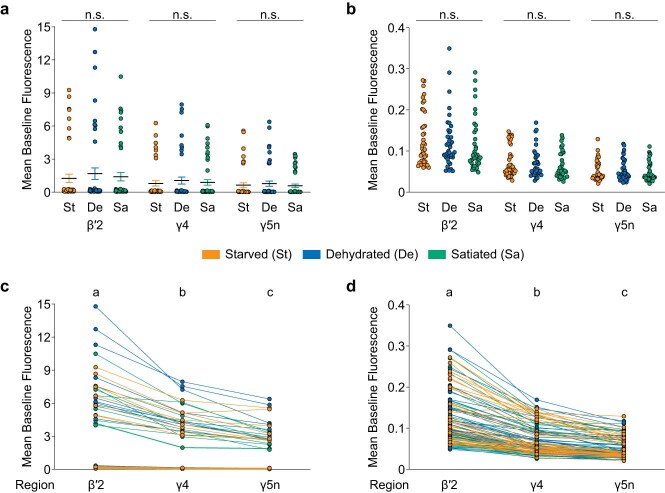
Resource-seeking behaviours are ordinarily constrained by physiological needs and threats of danger, and the loss of these controls is associated with pathological reward seeking1. Although dysfunction of the dopaminergic valuation system of the brain is known to contribute towards unconstrained reward seeking2,3, the underlying reasons for this behaviour are unclear. Here we describe dopaminergic neural mechanisms that produce reward seeking despite adverse consequences in Drosophila melanogaster. Odours paired with optogenetic activation of a defined subset of reward-encoding dopaminergic neurons become cues that starved flies seek while neglecting food and enduring electric shock punishment. Unconstrained seeking of reward is not observed after learning with sugar or synthetic engagement of other dopaminergic neuron populations. Antagonism between reward-encoding and punishment-encoding dopaminergic neurons accounts for the perseverance of reward seeking despite punishment, whereas synthetic engagement of the reward-encoding dopaminergic neurons also impairs the ordinary need-dependent dopaminergic valuation of available food. Connectome analyses reveal that the population of reward-encoding dopaminergic neurons receives highly heterogeneous input, consistent with parallel representation of diverse rewards, and recordings demonstrate state-specific gating and satiety-related signals. We propose that a similar dopaminergic valuation system dysfunction is likely to contribute to maladaptive seeking of rewards by mammals.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
Dopamine determines how reward overrides risk.Nature. 2023 Nov;623(7986):258-259. doi: 10.1038/d41586-023-03085-4. Nature. 2023. PMID: 37880521 Free PMC article.
References
-
- Diagnostic and Statistical Manual of Mental Disorders, 5th Edn (American Psychiatric Association, 2022).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
