

A single photoreceptor splits perception and entrainment by cotransmission
- PMID: 37880372
- PMCID: PMC10651484
- DOI: 10.1038/s41586-023-06681-6
A single photoreceptor splits perception and entrainment by cotransmission
Abstract
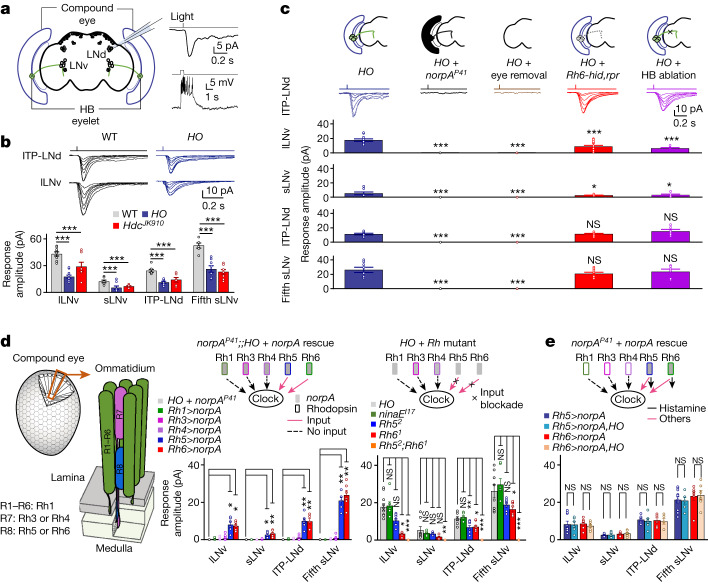
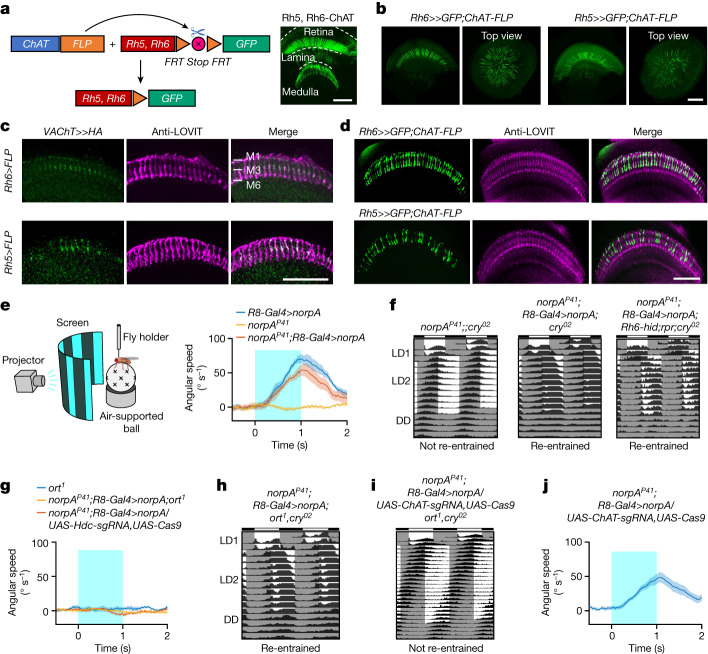
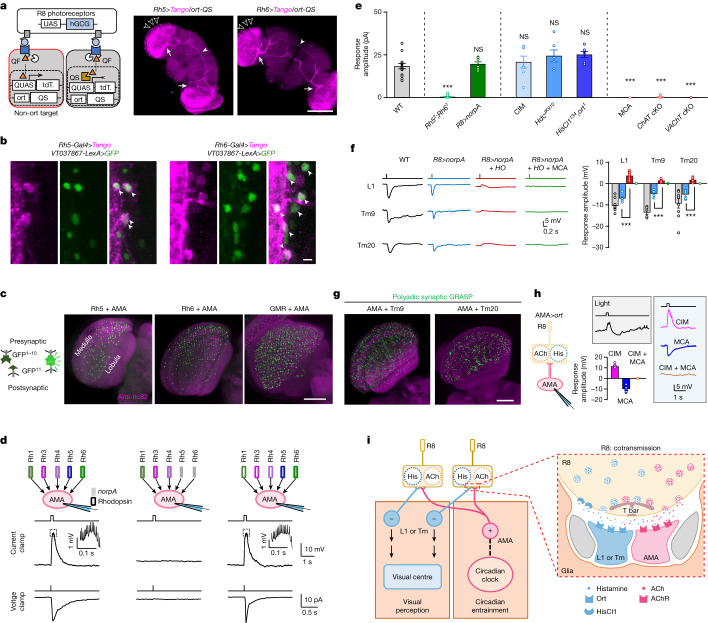
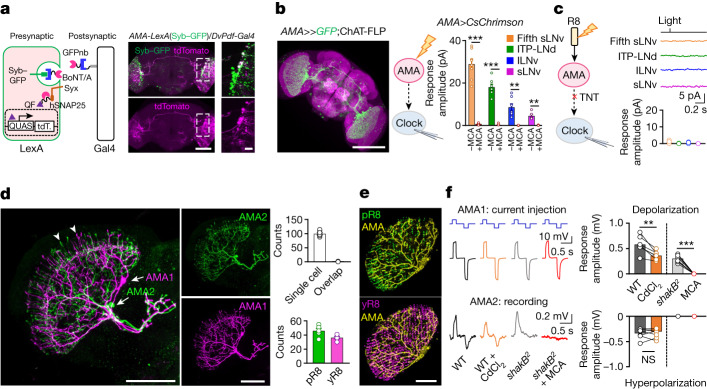
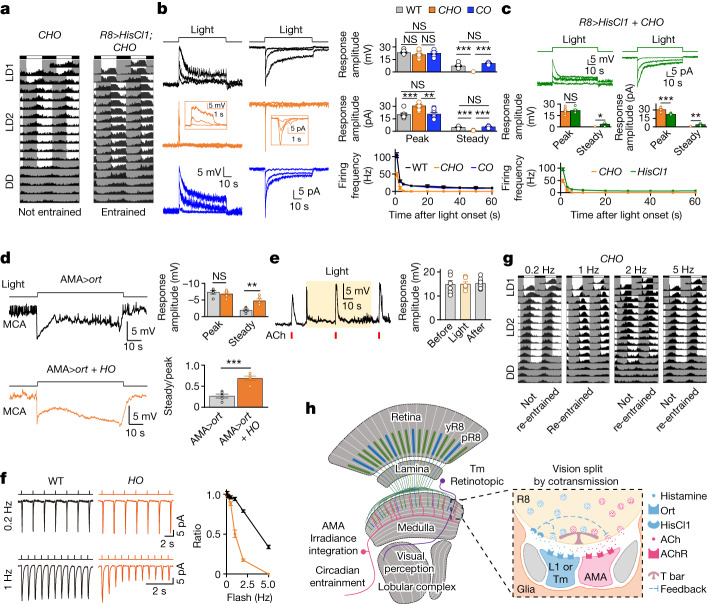
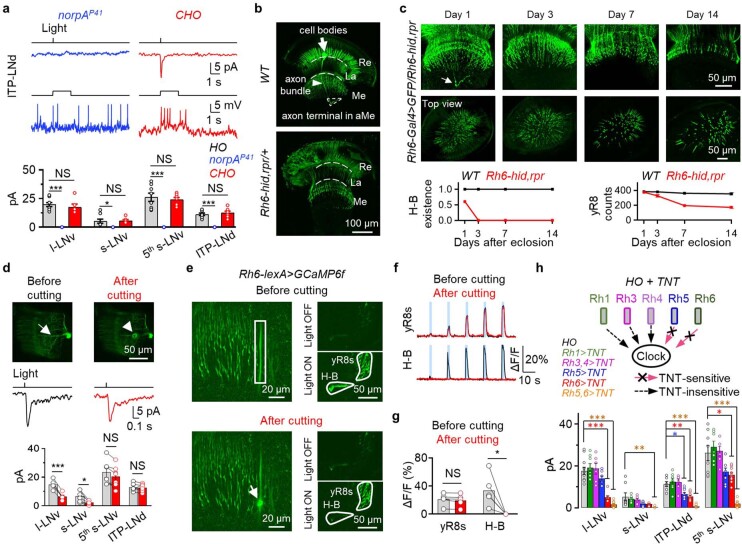
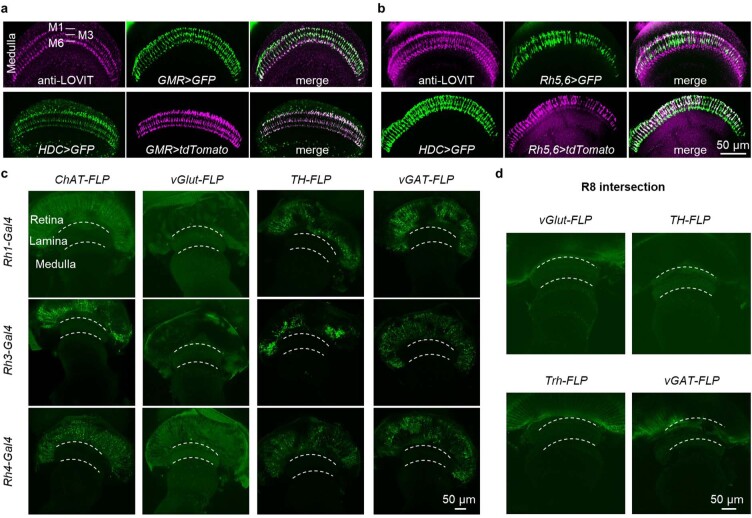
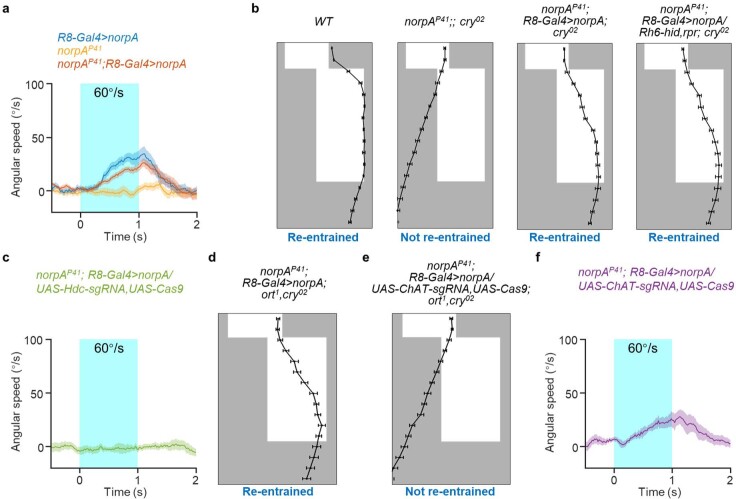
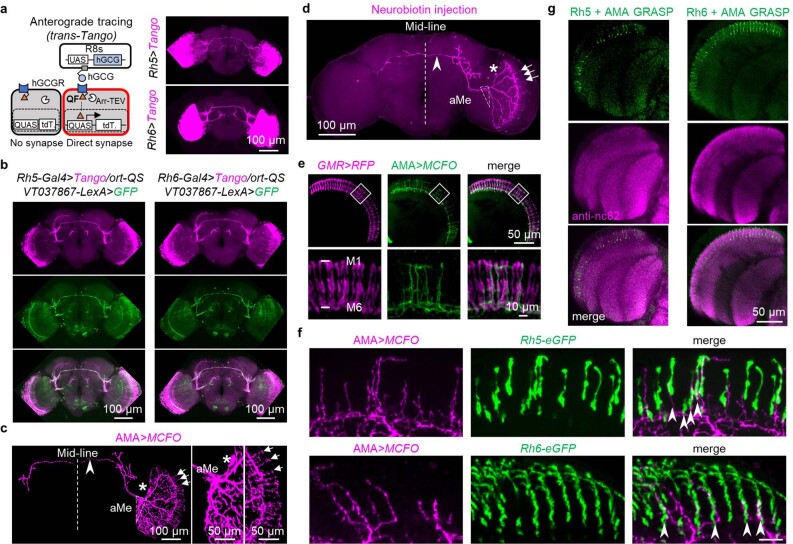
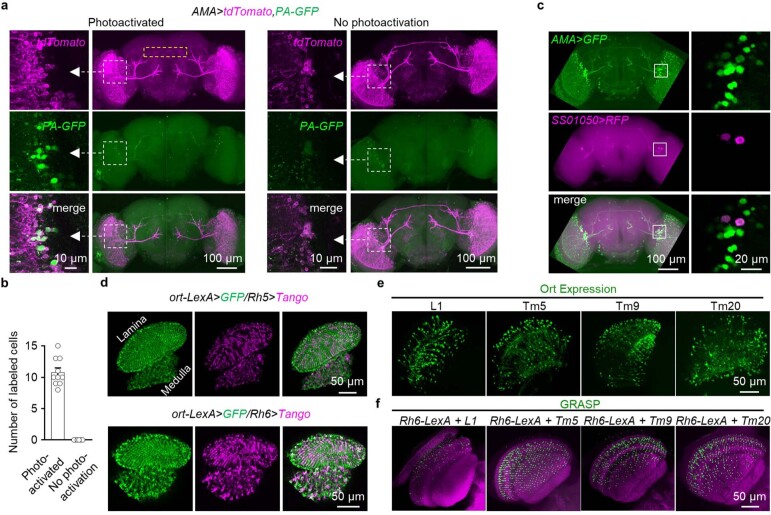
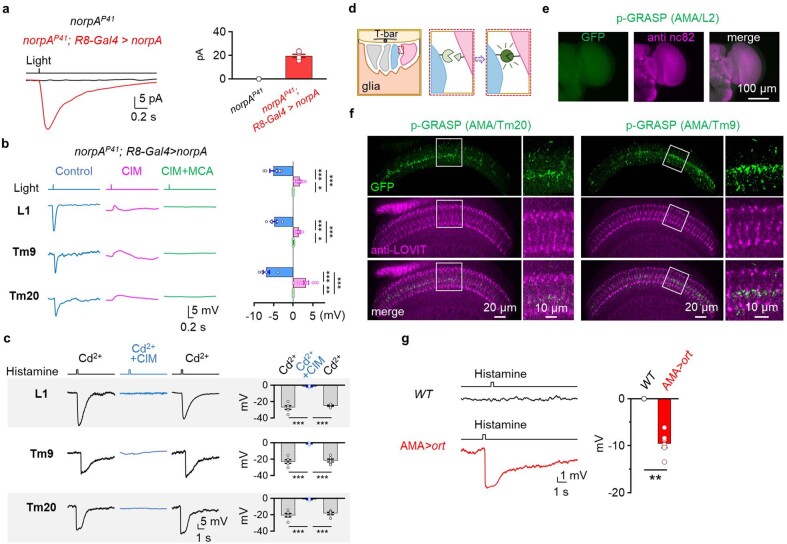
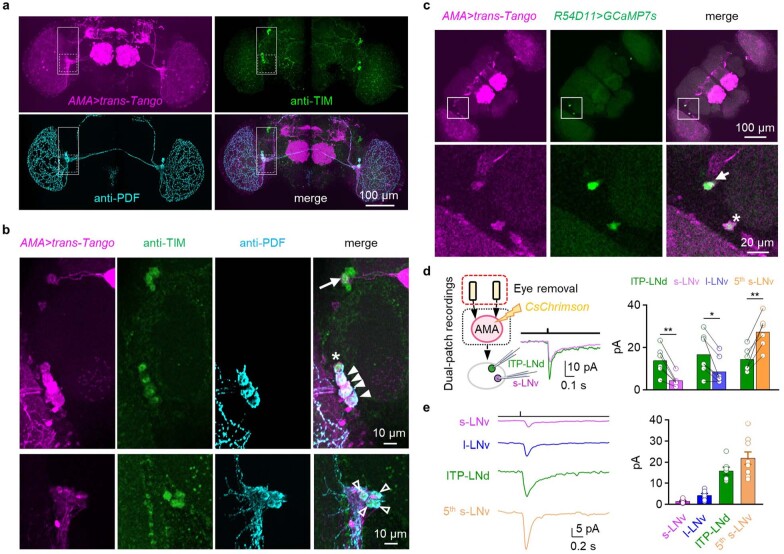
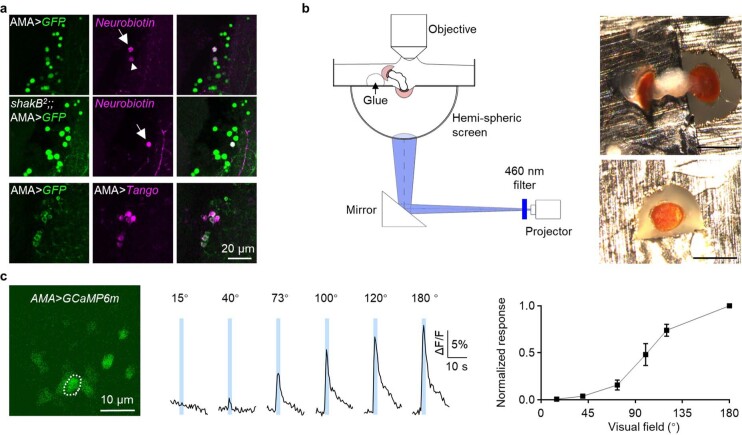
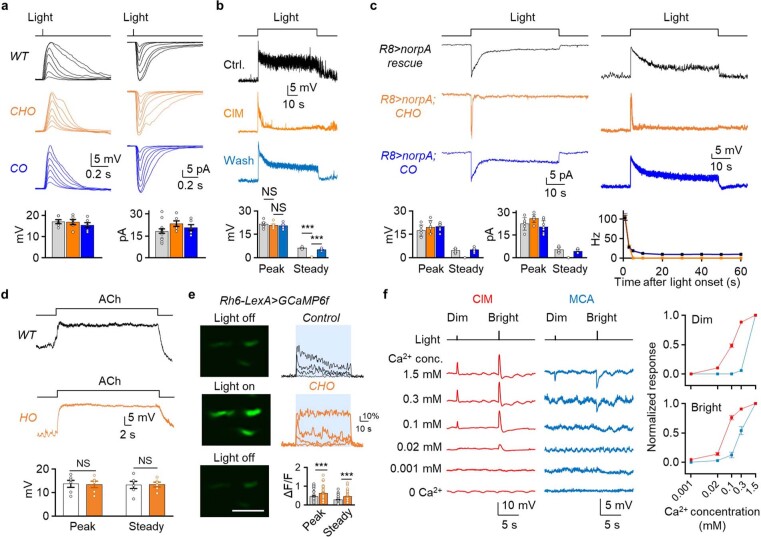
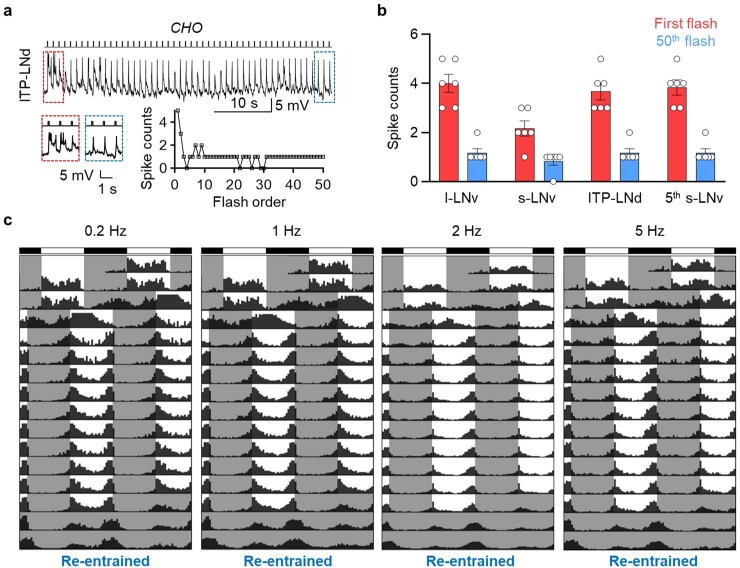
Vision enables both image-forming perception, driven by a contrast-based pathway, and unconscious non-image-forming circadian photoentrainment, driven by an irradiance-based pathway1,2. Although two distinct photoreceptor populations are specialized for each visual task3-6, image-forming photoreceptors can additionally contribute to photoentrainment of the circadian clock in different species7-15. However, it is unknown how the image-forming photoreceptor pathway can functionally implement the segregation of irradiance signals required for circadian photoentrainment from contrast signals required for image perception. Here we report that the Drosophila R8 photoreceptor separates image-forming and irradiance signals by co-transmitting two neurotransmitters, histamine and acetylcholine. This segregation is further established postsynaptically by histamine-receptor-expressing unicolumnar retinotopic neurons and acetylcholine-receptor-expressing multicolumnar integration neurons. The acetylcholine transmission from R8 photoreceptors is sustained by an autocrine negative feedback of the cotransmitted histamine during the light phase of light-dark cycles. At the behavioural level, elimination of histamine and acetylcholine transmission impairs R8-driven motion detection and circadian photoentrainment, respectively. Thus, a single type of photoreceptor can achieve the dichotomy of visual perception and circadian photoentrainment as early as the first visual synapses, revealing a simple yet robust mechanism to segregate and translate distinct sensory features into different animal behaviours.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
