

Diversity at single nucleotide to pangenome scales among sulfur cycling bacteria in salt marshes
- PMID: 37882526
- PMCID: PMC10686091
- DOI: 10.1128/aem.00988-23
Diversity at single nucleotide to pangenome scales among sulfur cycling bacteria in salt marshes
Abstract
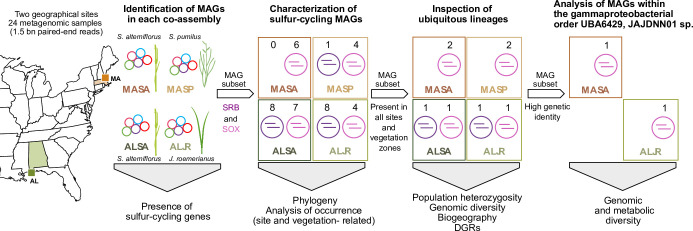
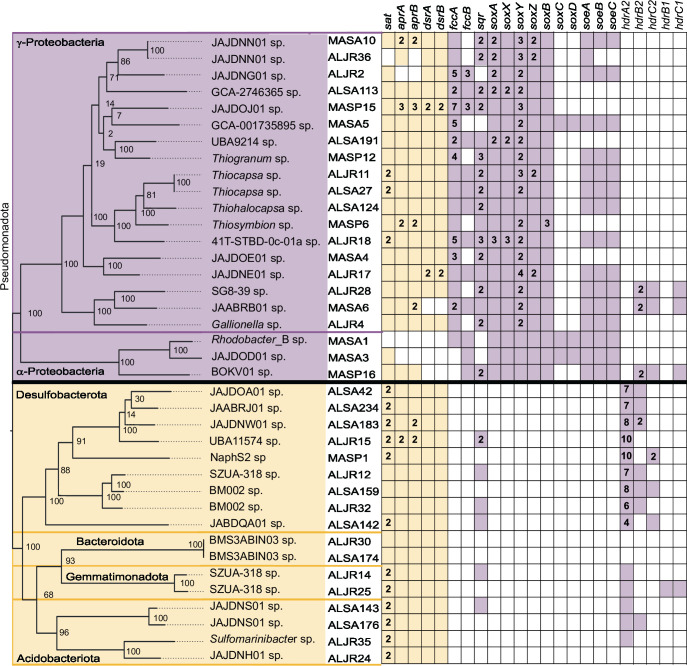
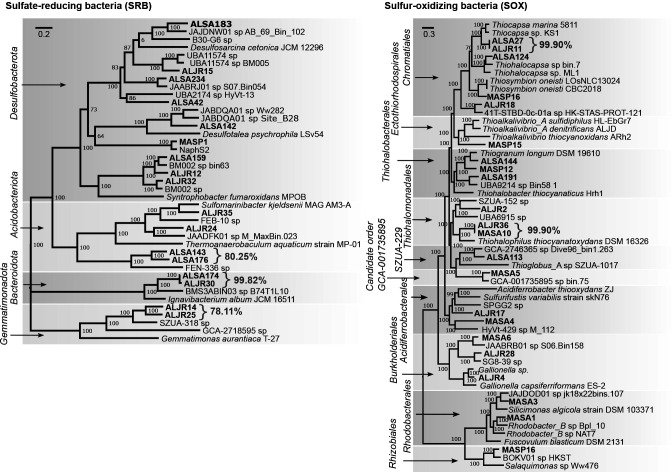
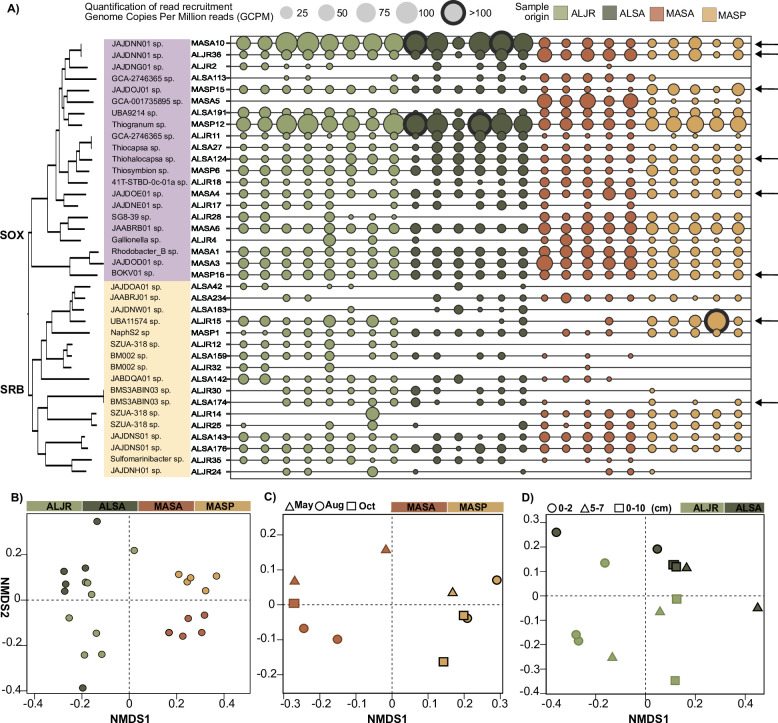
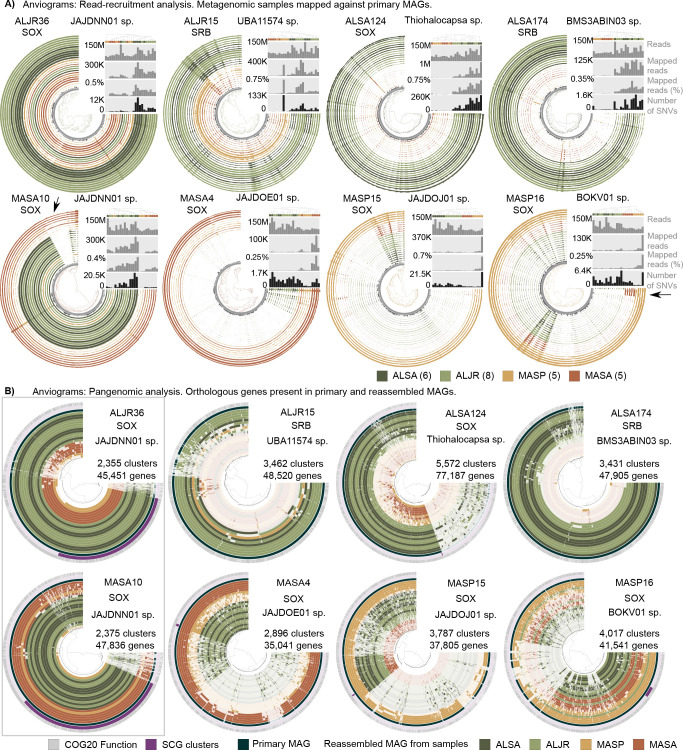
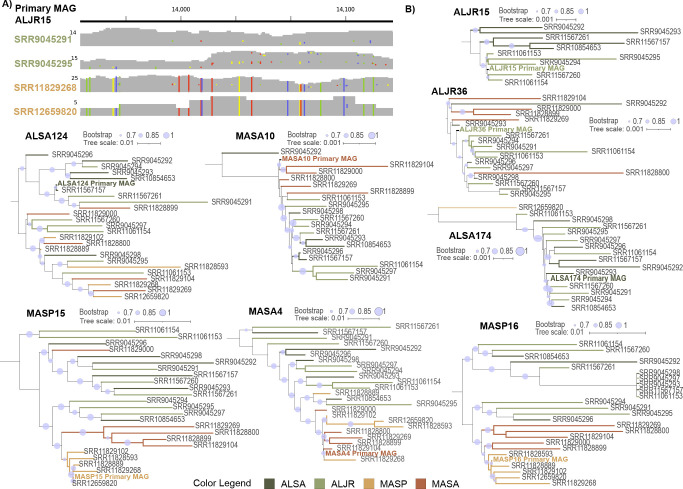
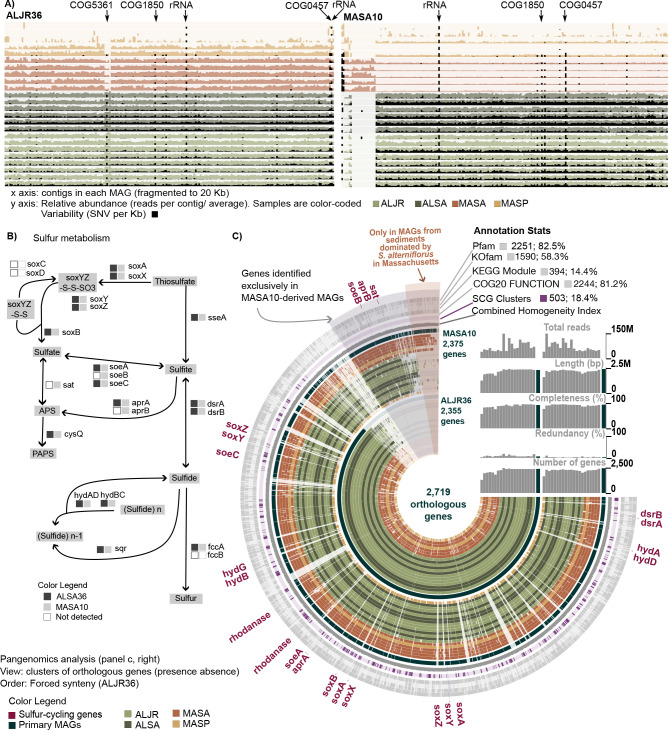
Salt marshes are known for their significant carbon storage capacity, and sulfur cycling is closely linked with the ecosystem-scale carbon cycling in these ecosystems. Sulfate reducers are key for the decomposition of organic matter, and sulfur oxidizers remove toxic sulfide, supporting the productivity of marsh plants. To date, the complexity of coastal environments, heterogeneity of the rhizosphere, high microbial diversity, and uncultured majority hindered our understanding of the genomic diversity of sulfur-cycling microbes in salt marshes. Here, we use comparative genomics to overcome these challenges and provide an in-depth characterization of sulfur-cycling microbial diversity in salt marshes. We characterize communities across distinct sites and plant species and uncover extensive genomic diversity at the taxon level and specific genomic features present in MAGs affiliated with uncultivated sulfur-cycling lineages. Our work provides insights into the partnerships in salt marshes and a roadmap for multiscale analyses of diversity in complex biological systems.
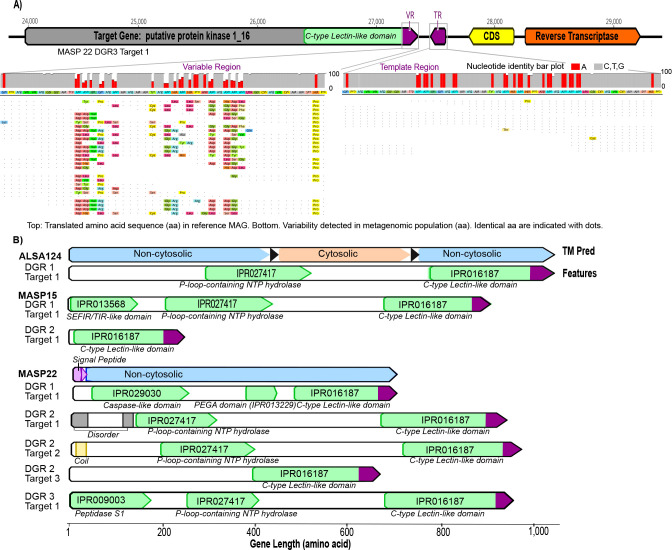
Keywords: diversity-generating retroelement; pangenomics; single-nucleotide polymorphism; site-specific genetic diversity; sulfate-reducing bacteria; sulfur-oxidizing bacteria.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Lee RW, Kraus DW, Doeller JE. 1999. Oxidation of sulfide by Spartina alterniflora roots. Limnol Oceanogr 44:1155–1159. doi: 10.4319/lo.1999.44.4.1155 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
