

This is a preprint.
Protein coopted from a phage restriction system dictates orthogonal cell division plane selection in Staphylococcus aureus
- PMID: 37886572
- PMCID: PMC10602043
- DOI: 10.1101/2023.09.03.556088
Protein coopted from a phage restriction system dictates orthogonal cell division plane selection in Staphylococcus aureus
Update in
-
PcdA promotes orthogonal division plane selection in Staphylococcus aureus.Nat Microbiol. 2024 Nov;9(11):2997-3012. doi: 10.1038/s41564-024-01821-8. Epub 2024 Oct 28. Nat Microbiol. 2024. PMID: 39468247 Free PMC article.
Abstract
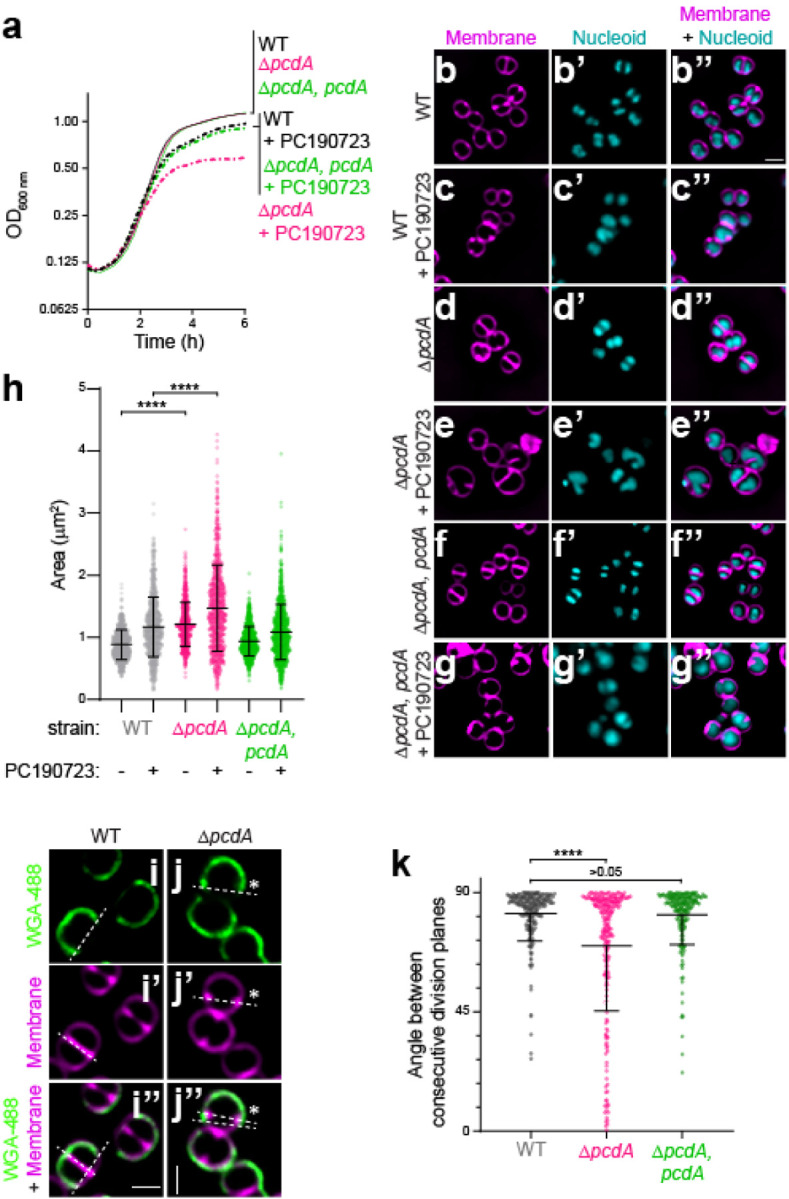
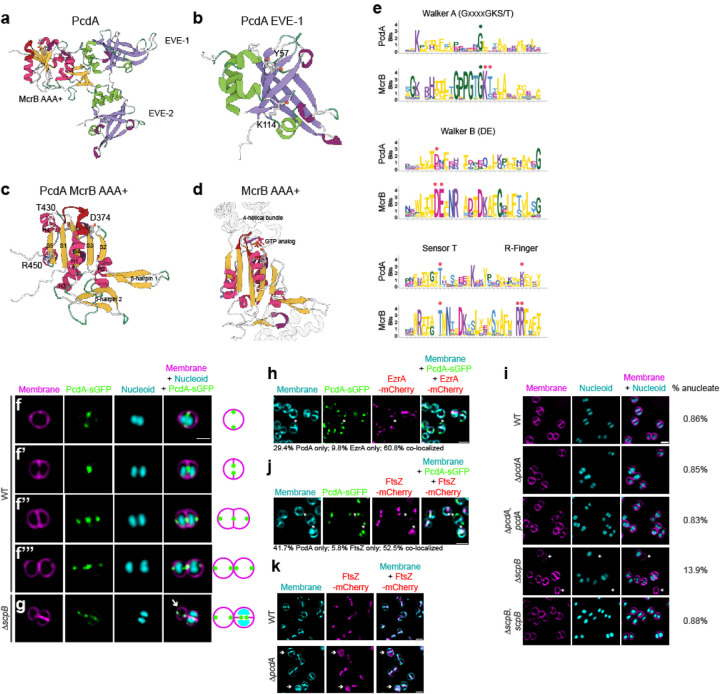
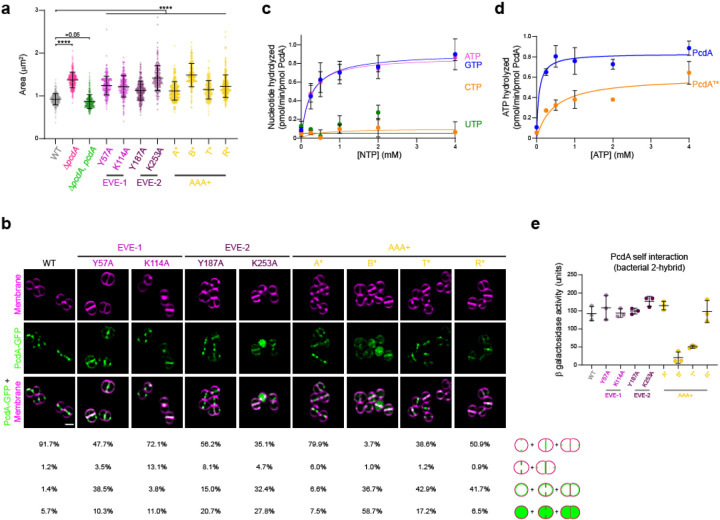
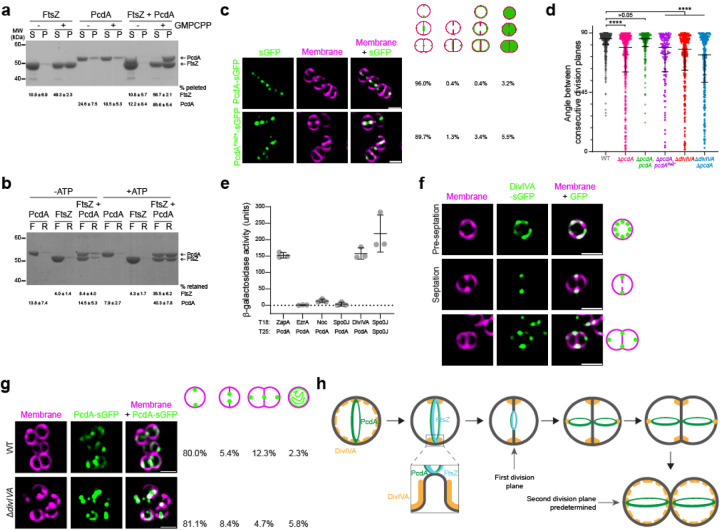
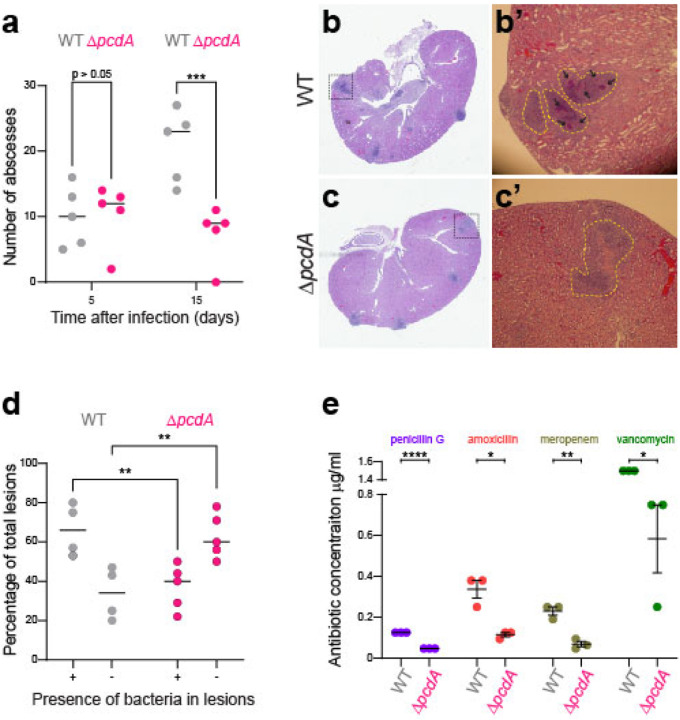
The spherical bacterium Staphylococcus aureus, a leading cause of nosocomial infections, undergoes binary fission by dividing in two alternating orthogonal planes, but the mechanism by which S. aureus correctly selects the next cell division plane is not known. To identify cell division placement factors, we performed a chemical genetic screen that revealed a gene which we termed pcdA. We show that PcdA is a member of the McrB family of AAA+ NTPases that has undergone structural changes and a concomitant functional shift from a restriction enzyme subunit to an early cell division protein. PcdA directly interacts with the tubulin-like central divisome component FtsZ and localizes to future cell division sites before membrane invagination initiates. This parallels the action of another McrB family protein, CTTNBP2, which stabilizes microtubules in animals. We show that PcdA also interacts with the structural protein DivIVA and propose that the DivIVA/PcdA complex recruits unpolymerized FtsZ to assemble along the proper cell division plane. Deletion of pcdA conferred abnormal, non-orthogonal division plane selection, increased sensitivity to cell wall-targeting antibiotics, and reduced virulence in a murine infection model. Targeting PcdA could therefore highlight a treatment strategy for combatting antibiotic-resistant strains of S. aureus.
Keywords: MRSA; Min system; MinCDE; MreB; vancomycin.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources