

Functionally comparable but evolutionarily distinct nucleotide-targeting effectors help identify conserved paradigms across diverse immune systems
- PMID: 37889040
- PMCID: PMC10681802
- DOI: 10.1093/nar/gkad879
Functionally comparable but evolutionarily distinct nucleotide-targeting effectors help identify conserved paradigms across diverse immune systems
Abstract
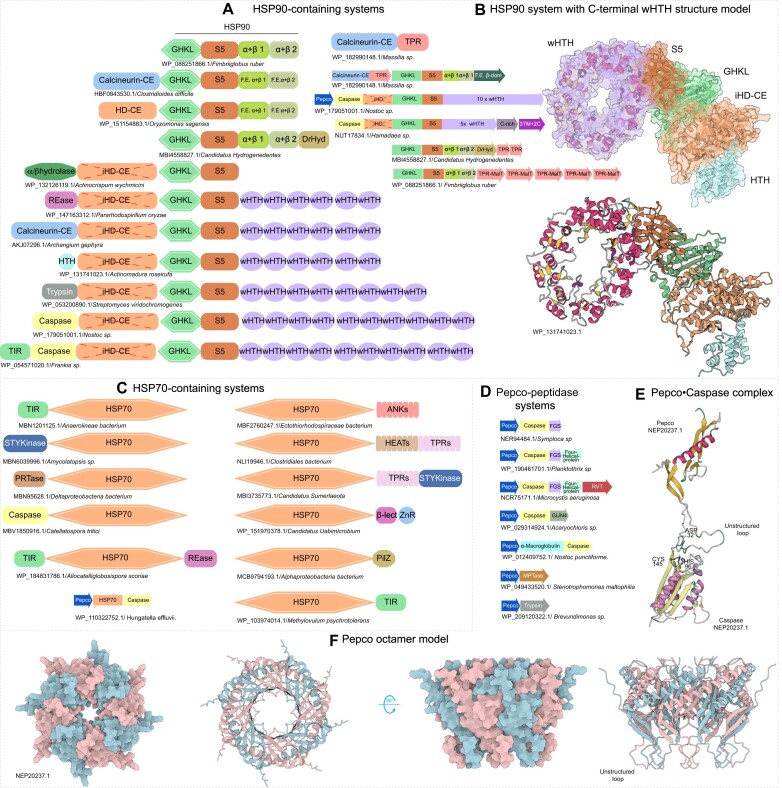
While nucleic acid-targeting effectors are known to be central to biological conflicts and anti-selfish element immunity, recent findings have revealed immune effectors that target their building blocks and the cellular energy currency-free nucleotides. Through comparative genomics and sequence-structure analysis, we identified several distinct effector domains, which we named Calcineurin-CE, HD-CE, and PRTase-CE. These domains, along with specific versions of the ParB and MazG domains, are widely present in diverse prokaryotic immune systems and are predicted to degrade nucleotides by targeting phosphate or glycosidic linkages. Our findings unveil multiple potential immune systems associated with at least 17 different functional themes featuring these effectors. Some of these systems sense modified DNA/nucleotides from phages or operate downstream of novel enzymes generating signaling nucleotides. We also uncovered a class of systems utilizing HSP90- and HSP70-related modules as analogs of STAND and GTPase domains that are coupled to these nucleotide-targeting- or proteolysis-induced complex-forming effectors. While widespread in bacteria, only a limited subset of nucleotide-targeting effectors was integrated into eukaryotic immune systems, suggesting barriers to interoperability across subcellular contexts. This work establishes nucleotide-degrading effectors as an emerging immune paradigm and traces their origins back to homologous domains in housekeeping systems.
Published by Oxford University Press on behalf of Nucleic Acids Research 2023.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
