

Microglial FoxO3a deficiency ameliorates ferroptosis-induced brain injury of intracerebral haemorrhage via regulating autophagy and heme oxygenase-1
- PMID: 37890842
- PMCID: PMC10805503
- DOI: 10.1111/jcmm.18007
Microglial FoxO3a deficiency ameliorates ferroptosis-induced brain injury of intracerebral haemorrhage via regulating autophagy and heme oxygenase-1
Abstract
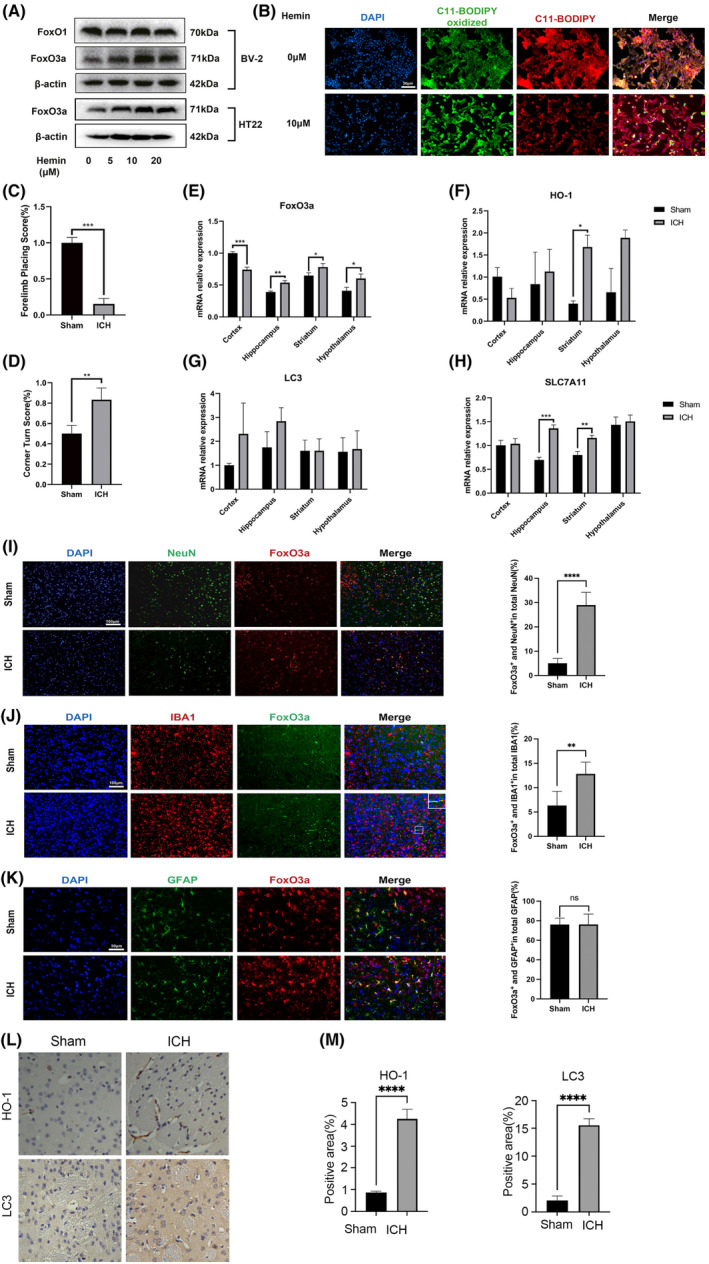
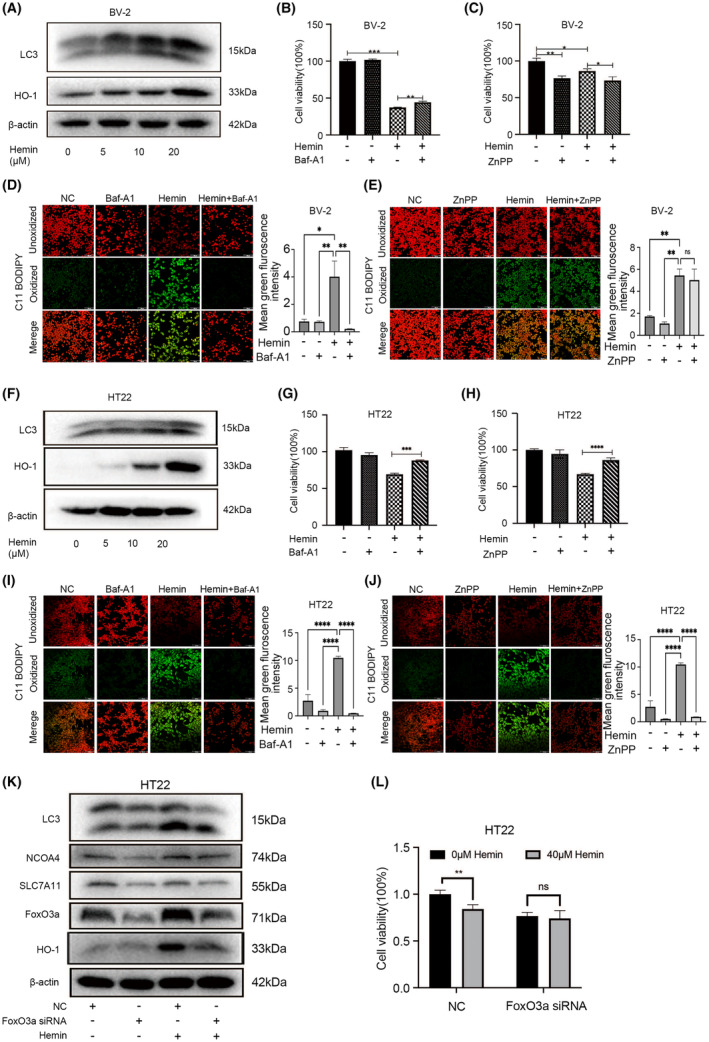
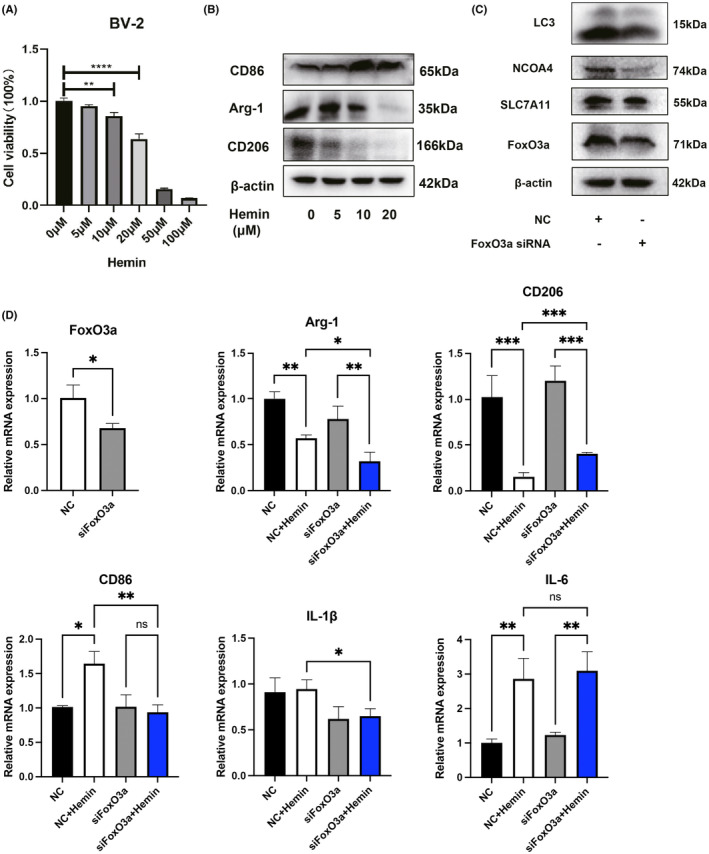
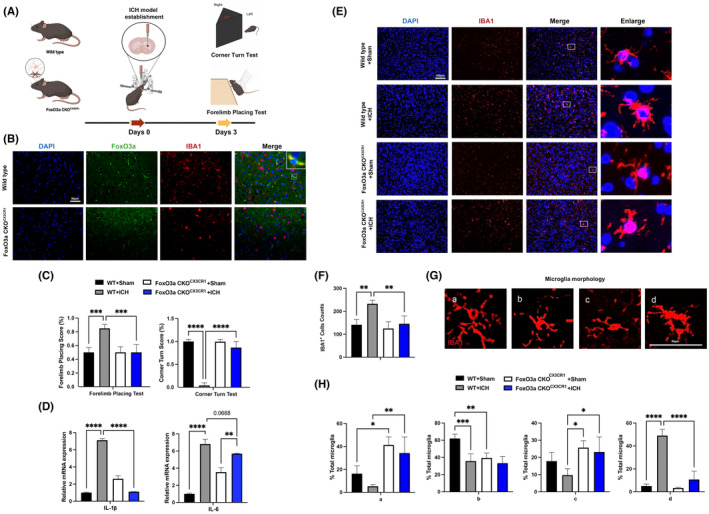
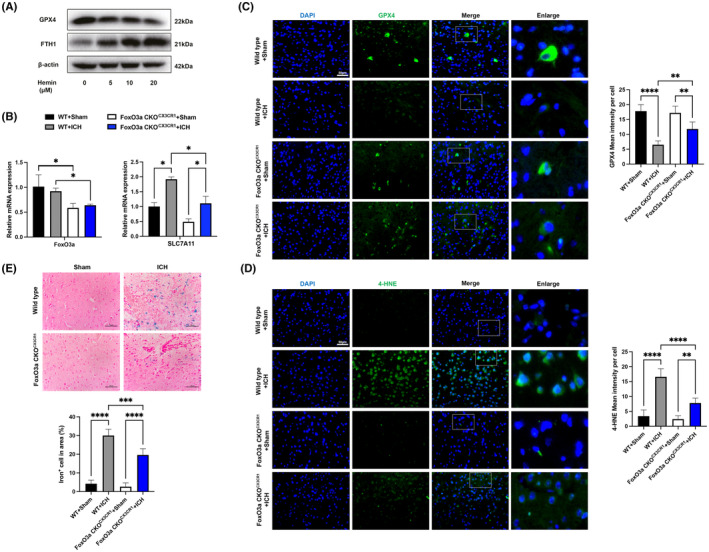
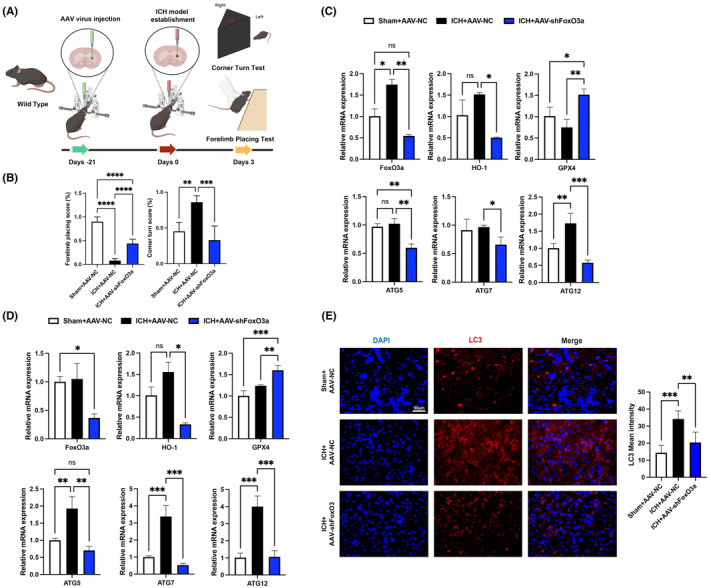
Microglial HO-1 regulates iron metabolism in the brain. Intracerebral haemorrhage (ICH) shares features of ferroptosis and necroptosis; hemin is an oxidized product of haemoglobin from lysed red blood cells, leading to secondary injury. However, little is known about the underlying molecular mechanisms attributable to secondary injury by hemin or ICH. In this study, we first show that FoxO3a was highly co-located with neurons and microglia but not astrocytes area of ICH model mice. Hemin activated FoxO3a/ATG-mediated autophagy and HO-1 signalling resulting in ferroptosis in vitro and in a mice model of brain haemorrhage. Accordingly, autophagy inhibitor Baf-A1 or HO-1 inhibitor ZnPP protected against hemin-induced ferroptosis. Hemin promoted ferroptosis of neuronal cells via FoxO3a/ATG-mediated autophagy and HO-1 signalling pathway. Knock-down of FoxO3a inhibited autophagy and prevented hemin-induced ferroptosis dependent of HO-1 signalling. We first showed that hemin stimulated microglial FoxO3a/HO-1 expression and enhanced the microglial polarisation towards the M1 phenotype, while knockdown of microglial FoxO3a inhibited pro-inflammatory cytokine production in microglia. Furthermore, the microglia activation in the striatum showed significant along with a high expression level of FoxO3a in the ICH mice. We found that conditional knockout of FoxO3a in microglia in mice alleviated neurological deficits and microglia activation as well as ferroptosis-induced striatum injury in the autologous blood-induced ICH model. We demonstrate, for the first time, that FoxO3a/ATG-mediated autophagy and HO-1 play an important role in microglial activation and ferroptosis-induced striatum injury of ICH, identifying a new therapeutic avenue for the treatment of ICH.
Keywords: FoxO3a; HO-1; ferroptosis; intracerebral haemorrhage; microglia.
© 2023 The Authors. Journal of Cellular and Molecular Medicine published by Foundation for Cellular and Molecular Medicine and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Yang Z, Liu B, Zhong L, et al. Toll‐like receptor‐4‐mediated autophagy contributes to microglial activation and inflammatory injury in mouse models of intracerebral haemorrhage. Neuropathol Appl Neurobiol. 2015;41(4):e95‐e106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
