

The regulatory subunits of CK2 complex mediate DNA damage response and virulence in Candida Glabrata
- PMID: 37891489
- PMCID: PMC10612253
- DOI: 10.1186/s12866-023-03069-4
The regulatory subunits of CK2 complex mediate DNA damage response and virulence in Candida Glabrata
Abstract
Background: Candida glabrata which belongs to normal microbiota, has caused significant concern worldwide due to its high prevalence and drug resistance in recent years. C. glabrata has developed many strategies to evade the clearance of the host immune system, thereby causing persistent infection. Although coping with the induced DNA damage is widely acknowledged to be important, the underlying mechanisms remain unclear.
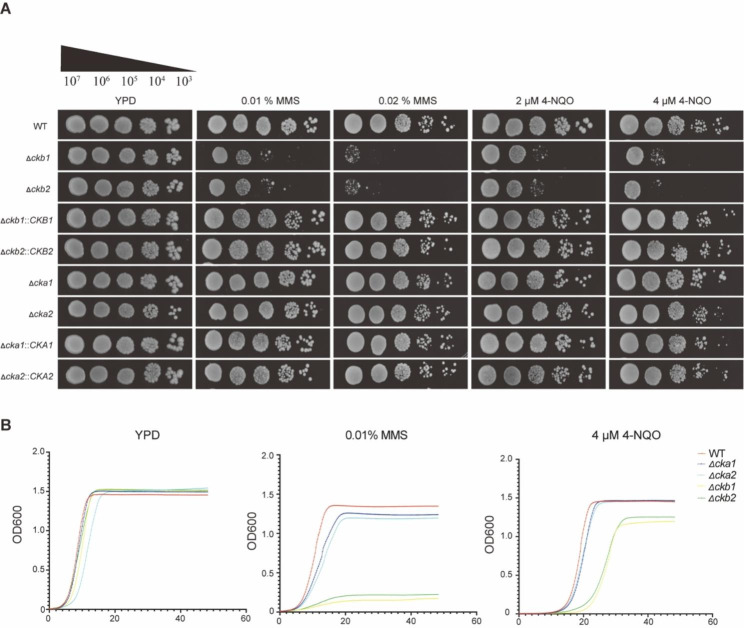
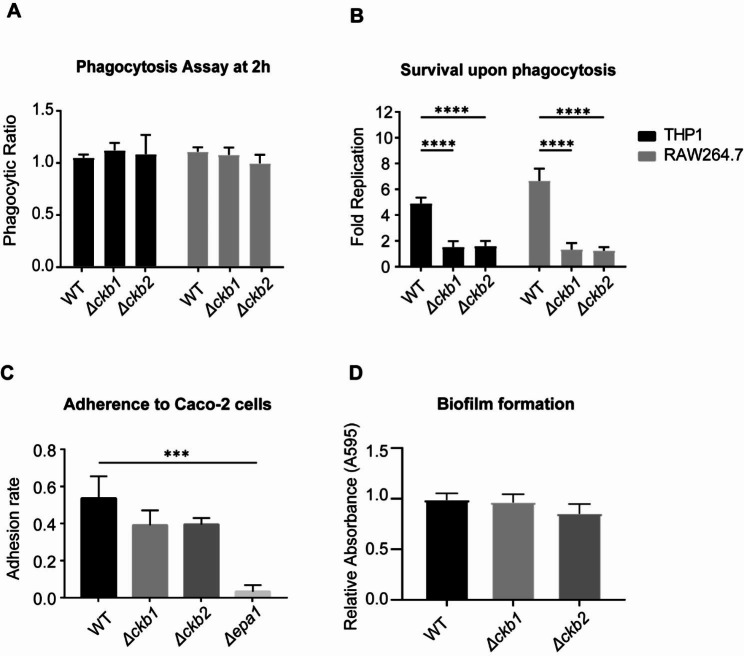
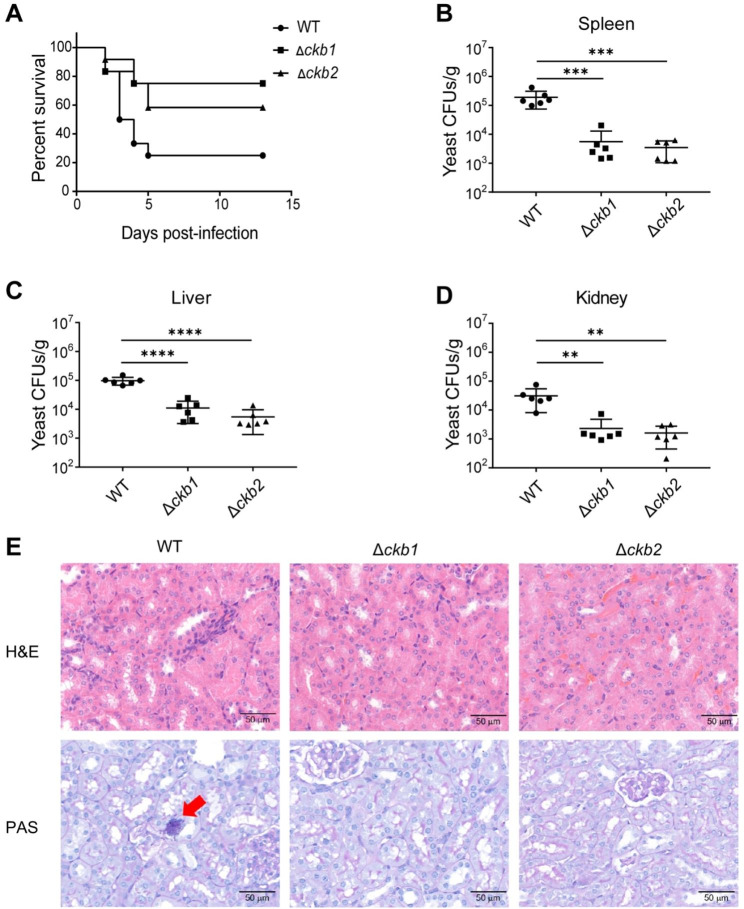
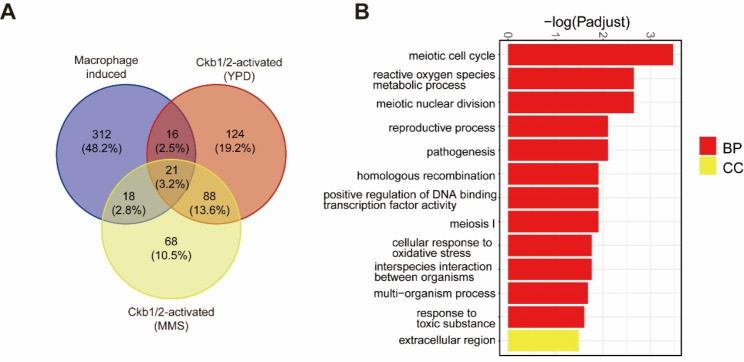
Results: The present study provides hitherto undocumented evidence of the importance of the regulatory subunits of CgCK2 (CgCkb1 and CgCkb2) in response to DNA damage. Deletion of CgCKB1 or CgCKB2 enhanced cellular apoptosis and DNA breaks and led to cell cycle delay. In addition, deficiencies in survival upon phagocytosis were observed in Δckb1 and Δckb2 strains. Consistently, disruption of CgCKB1 and CgCKB2 attenuated the virulence of C. glabrata in mouse models of invasive candidiasis. Furthermore, global transcriptional profiling analysis revealed that CgCkb1 and CgCkb2 participate in cell cycle resumption and genomic stability.
Conclusions: Overall, our findings suggest that the response to DNA damage stress is crucial for C. glabrata to survive in macrophages, leading to full virulence in vivo. The significance of this work lies in providing a better understanding of pathogenicity in C. glabrata-related candidiasis and expanding ideas for clinical therapies.
Keywords: Candida glabrata; Cell cycle; DNA damage; Macrophage; Virulence.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
