

Functional Modules in the Meristems: "Tinkering" in Action
- PMID: 37896124
- PMCID: PMC10610496
- DOI: 10.3390/plants12203661
Functional Modules in the Meristems: "Tinkering" in Action
Abstract
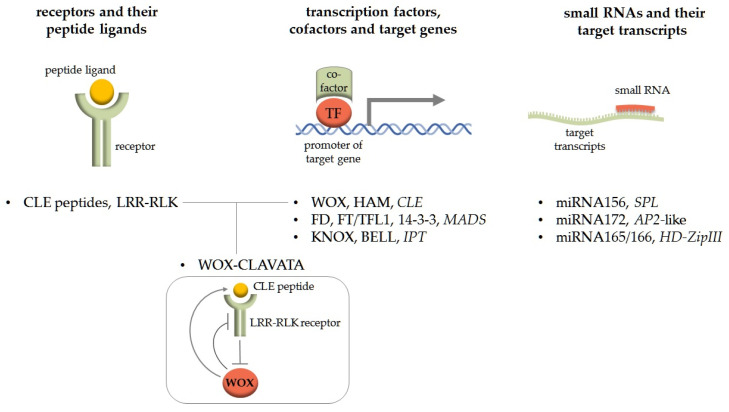
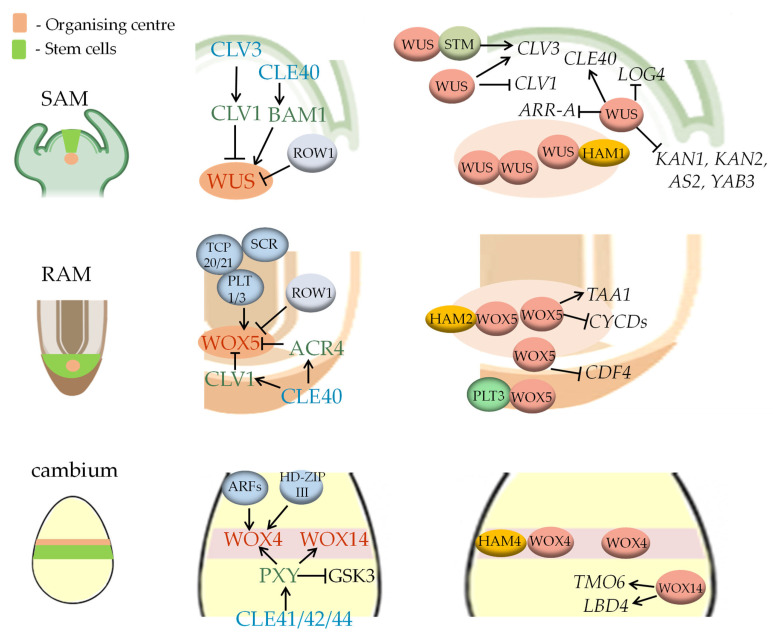
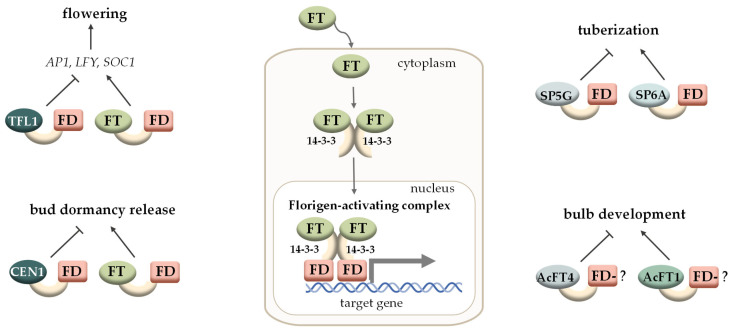
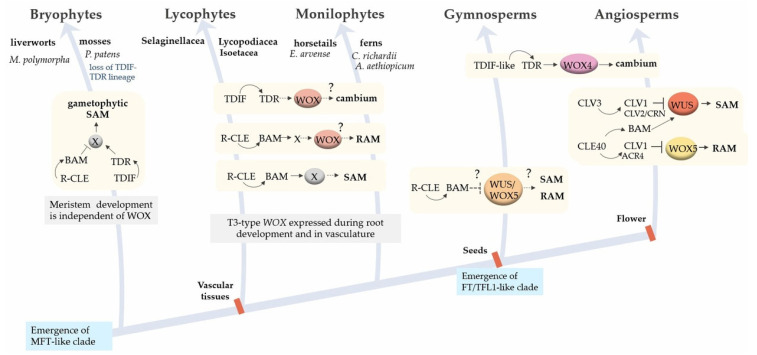
Background: A feature of higher plants is the modular principle of body organisation. One of these conservative morphological modules that regulate plant growth, histogenesis and organogenesis is meristems-structures that contain pools of stem cells and are generally organised according to a common principle. Basic content: The development of meristems is under the regulation of molecular modules that contain conservative interacting components and modulate the expression of target genes depending on the developmental context. In this review, we focus on two molecular modules that act in different types of meristems. The WOX-CLAVATA module, which includes the peptide ligand, its receptor and the target transcription factor, is responsible for the formation and control of the activity of all meristem types studied, but it has its own peculiarities in different meristems. Another regulatory module is the so-called florigen-activated complex, which is responsible for the phase transition in the shoot vegetative meristem (e.g., from the vegetative shoot apical meristem to the inflorescence meristem).
Conclusions: The review considers the composition and functions of these two functional modules in different developmental programmes, as well as their appearance, evolution and use in plant breeding.
Keywords: evo-devo; functional module; land plants; meristem; peptide/protein ligand; phase transition; regulation of transcription.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Notov A.A. On Functional Organization and Individual Development of Modular Objects. Wulfenia. 2005;12:65–85.
-
- Klingenberg C.P. Morphological Integration and Developmental Modularity. Annu. Rev. Ecol. Evol. Syst. 2008;39:115–132. doi: 10.1146/annurev.ecolsys.37.091305.110054. - DOI
-
- Afonkin S.Y. Biokombinatorika Ili Blochnyi Printsyp Organizasii Zhizni [Bio-Combination Theory or Block Principle of the Organisation of a Life] Biology. 2003;35
-
- Inge-Vechtomov S.G. Fundamental’nye Zoologicheskie Issledovaniya (Fundamental Zoological Studies) KMK; St. Petersburg, Russia: 2004. Block Principle in the Theory of Evolution. Prospects and Paradoxes; pp. 74–88.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
