

This is a preprint.
High-density recording reveals sparse clusters (but not columns) for shape and texture encoding in macaque V4
- PMID: 37904996
- PMCID: PMC10614825
- DOI: 10.1101/2023.10.15.562424
High-density recording reveals sparse clusters (but not columns) for shape and texture encoding in macaque V4
Update in
-
High-Density Recording Reveals Sparse Clusters (But Not Columns) for Shape and Texture Encoding in Macaque V4.J Neurosci. 2025 Jan 29;45(5):e1893232024. doi: 10.1523/JNEUROSCI.1893-23.2024. J Neurosci. 2025. PMID: 39562041 Free PMC article.
Abstract
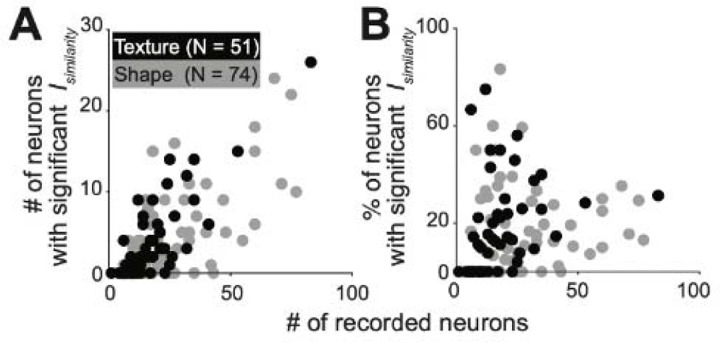
Macaque area V4 includes neurons that exhibit exquisite selectivity for visual form and surface texture, but their functional organization across laminae is unknown. We used high-density Neuropixels probes in two awake monkeys to characterize shape and texture tuning of dozens of neurons simultaneously across layers. We found sporadic clusters of neurons that exhibit similar tuning for shape and texture: ~20% exhibited similar tuning with their neighbors. Importantly, these clusters were confined to a few layers, seldom 'columnar' in structure. This was the case even when neurons were strongly driven, and exhibited robust contrast invariance for shape and texture tuning. We conclude that functional organization in area V4 is not columnar for shape and texture stimulus features and in general organization maybe at a coarse scale (e.g. encoding of 2D vs 3D shape) rather than at a fine scale in terms of similarity in tuning for specific features (as in the orientation columns in V1). We speculate that this may be a direct consequence of the great diversity of inputs integrated by V4 neurons to build variegated tuning manifolds in a high-dimensional space.
Conflict of interest statement
Declaration of interests The authors declare that they have no competing interests.
Figures









References
-
- Albright TD, Desimone R, Gross CG (1984) Columnar organization of directionally selective cells in visual area MT of the macaque. J Neurophysiol 51, 16–31. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources