

This is a preprint.
Fine-grained descending control of steering in walking Drosophila
- PMID: 37904997
- PMCID: PMC10614758
- DOI: 10.1101/2023.10.15.562426
Fine-grained descending control of steering in walking Drosophila
Update in
-
Fine-grained descending control of steering in walking Drosophila.Cell. 2024 Oct 31;187(22):6290-6308.e27. doi: 10.1016/j.cell.2024.08.033. Epub 2024 Sep 17. Cell. 2024. PMID: 39293446 Free PMC article.
Abstract
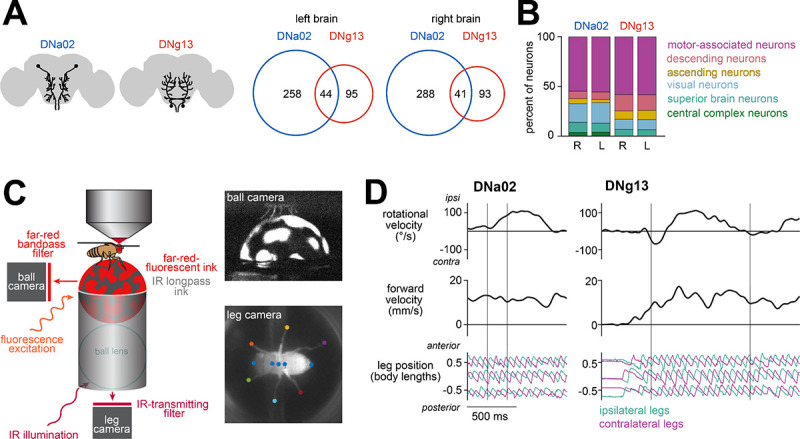
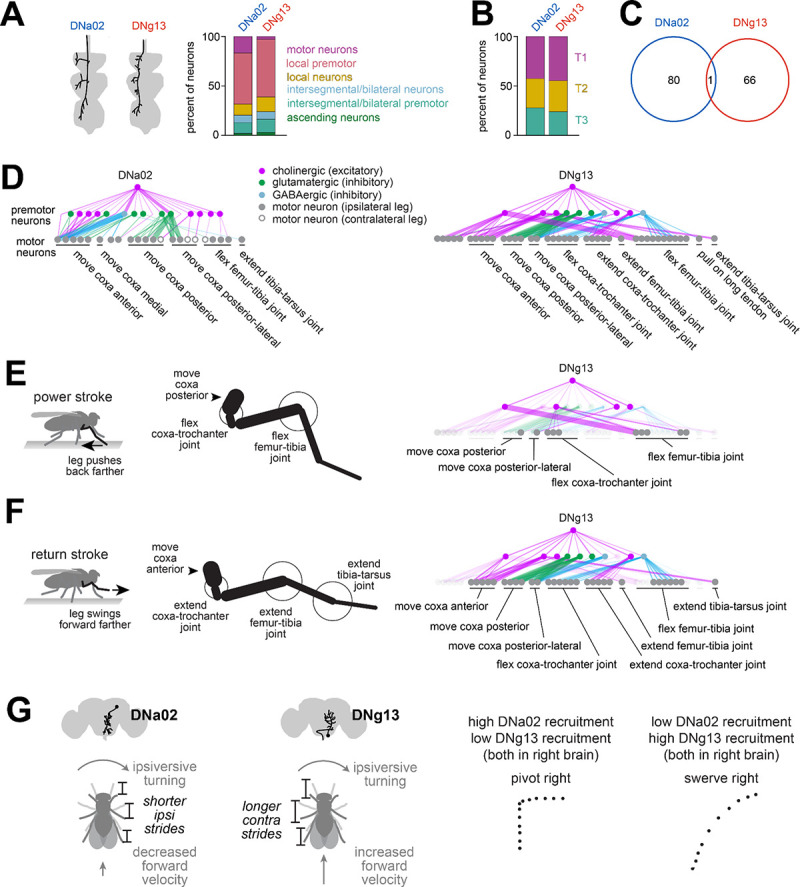
Locomotion involves rhythmic limb movement patterns that originate in circuits outside the brain. Purposeful locomotion requires descending commands from the brain, but we do not understand how these commands are structured. Here we investigate this issue, focusing on the control of steering in walking Drosophila. First, we describe different limb "gestures" associated with different steering maneuvers. Next, we identify a set of descending neurons whose activity predicts steering. Focusing on two descending cell types downstream from distinct brain networks, we show that they evoke specific limb gestures: one lengthens strides on the outside of a turn, while the other attenuates strides on the inside of a turn. Notably, a single descending neuron can have opposite effects during different locomotor rhythm phases, and we identify networks positioned to implement this phase-specific gating. Together, our results show how purposeful locomotion emerges from brain cells that drive specific, coordinated modulations of low-level patterns.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





References
-
- Brown T.G., and Sherrington C.S. (1997). The intrinsic factors in the act of progression in the mammal. Proc. R. Soc. Lond. B Biol. Sci. 84, 308–319.
-
- Grillner S. (2011). Control of locomotion in bipeds, tetrapods, and fish. In Handbook of Physiology: The Nervous System: Motor Control (John Wiley & Sons, Inc.).
-
- Graham D. (1979). Effects of circum-oesophageal lesion on the behaviour of the stick insect Carausius morosus. Biol. Cybern. 32, 139–145.
-
- Ryckebusch S., and Laurent G. (1993). Rhythmic patterns evoked in locust leg motor neurons by the muscarinic agonist pilocarpine. J. Neurophysiol. 69, 1583–1595. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources