

This is a preprint.
Skeletal muscle BMAL1 is necessary for transcriptional adaptation of local and peripheral tissues in response to endurance exercise training
- PMID: 37905004
- PMCID: PMC10614785
- DOI: 10.1101/2023.10.13.562100
Skeletal muscle BMAL1 is necessary for transcriptional adaptation of local and peripheral tissues in response to endurance exercise training
Update in
-
Skeletal muscle BMAL1 is necessary for transcriptional adaptation of local and peripheral tissues in response to endurance exercise training.Mol Metab. 2024 Aug;86:101980. doi: 10.1016/j.molmet.2024.101980. Epub 2024 Jun 29. Mol Metab. 2024. PMID: 38950777 Free PMC article.
Abstract
Objectives: In this investigation, we addressed the contribution of the core circadian clock factor, BMAL1, in skeletal muscle to both acute transcriptional responses to exercise and transcriptional remodelling in response to exercise training. Additionally, we adopted a systems biology approach to investigate how loss of skeletal muscle BMAL1 altered peripheral tissue homeostasis as well as exercise training adaptations in iWAT, liver, heart, and lung of male mice.
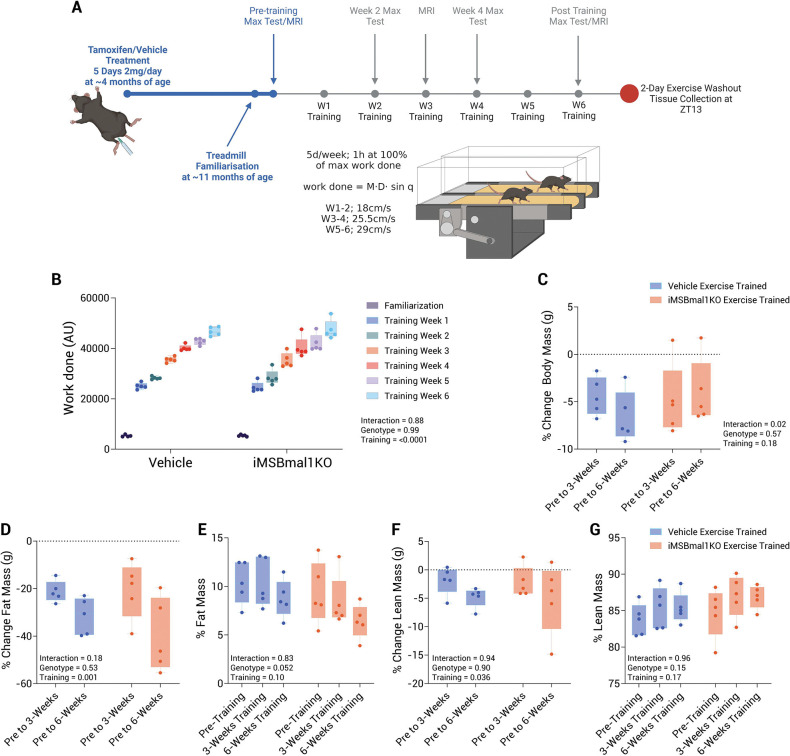
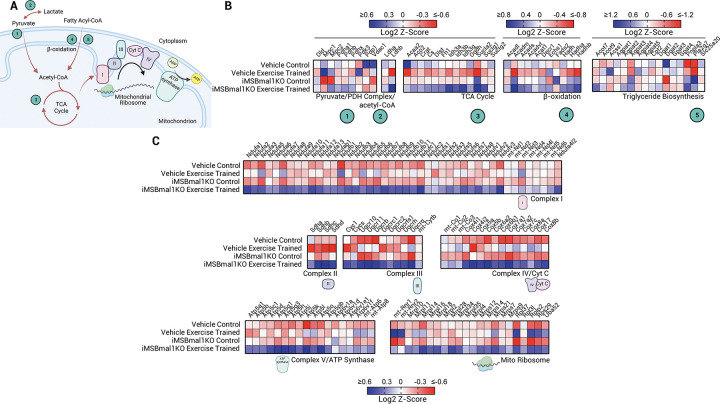
Methods: Combining inducible skeletal muscle specific BMAL1 knockout mice, physiological testing and standardized exercise protocols, we performed a multi-omic analysis (transcriptomics, chromatin accessibility and metabolomics) to explore loss of muscle BMAL1 on muscle and peripheral tissue responses to exercise.
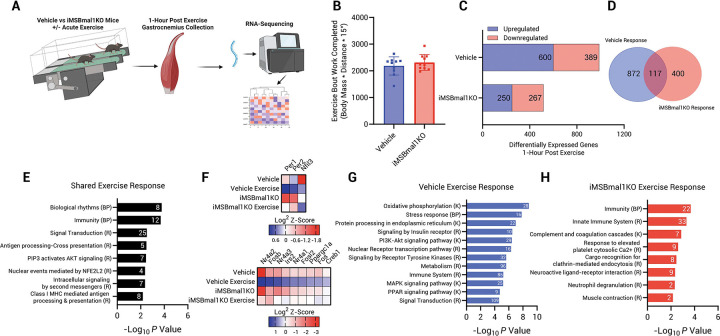
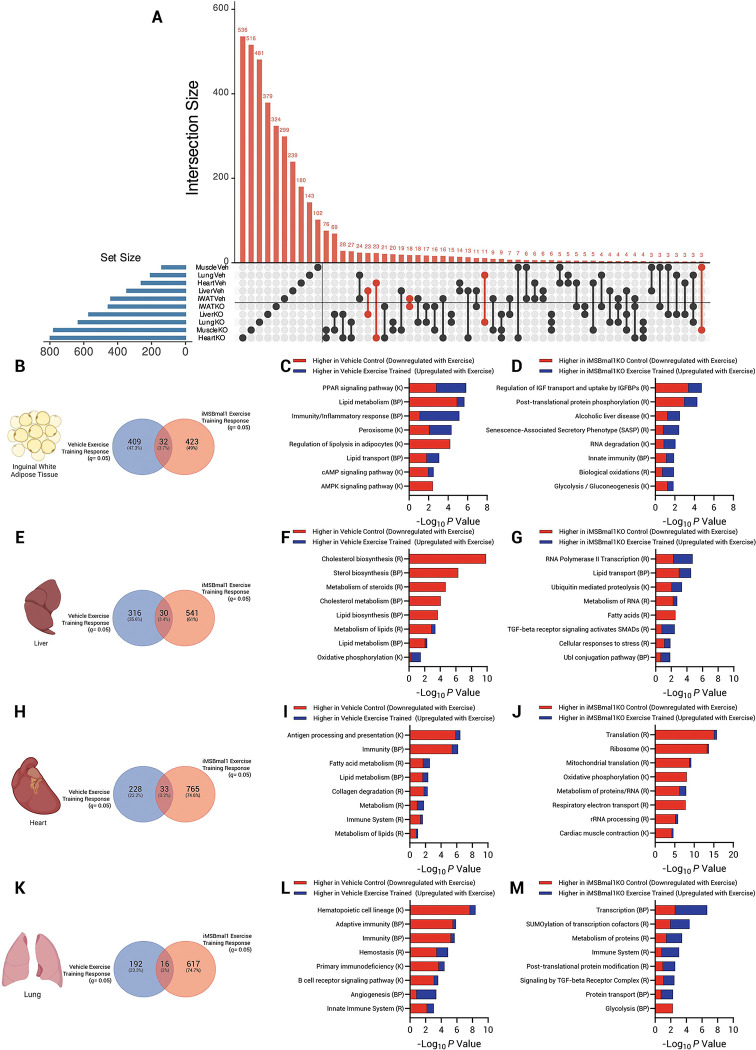
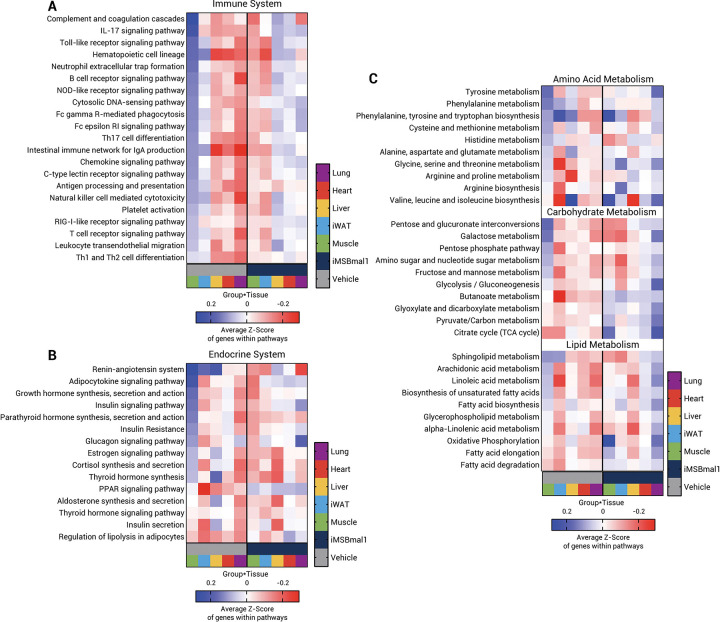
Results: Muscle-specific BMAL1 knockout mice demonstrated a blunted transcriptional response to acute exercise, characterized by the lack of upregulation of well-established exercise responsive transcription factors including Nr4a3 and Ppargc1a. Six weeks of exercise training in muscle-specific BMAL1 knockout mice induced significantly greater and divergent transcriptomic and metabolomic changes in muscle. Surprisingly, liver, lung, inguinal white adipose and heart showed divergent exercise training transcriptomes with less than 5% of 'exercise-training' responsive genes shared for each tissue between genotypes.
Conclusion: Our investigation has uncovered the critical role that BMAL1 plays in skeletal muscle as a key regulator of gene expression programs for both acute exercise and training adaptations. In addition, our work has uncovered the significant impact that altered exercise response in muscle plays in the peripheral tissue adaptation to exercise training. We also note that the transcriptome adaptations to steady state training suggest that without BMAL1, skeletal muscle does not achieve the expected homeostatic program. Our work also demonstrates that if the muscle adaptations diverge to a more maladaptive state this is linked to increased inflammation across many tissues. Understanding the molecular targets and pathways contributing to health vs. maladaptive exercise adaptations will be critical for the next stage of therapeutic design for exercise mimetics.
Keywords: Circadian Biology; Exercise; Inflammation; Metabolism; Signal Transduction; Transcription; circadian; skeletal muscle.
Conflict of interest statement
Declaration of Interests: The authors declare no competing interests.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources