

PFTK1 kinase regulates axogenesis during development via RhoA activation
- PMID: 37907898
- PMCID: PMC10617079
- DOI: 10.1186/s12915-023-01732-w
PFTK1 kinase regulates axogenesis during development via RhoA activation
Abstract
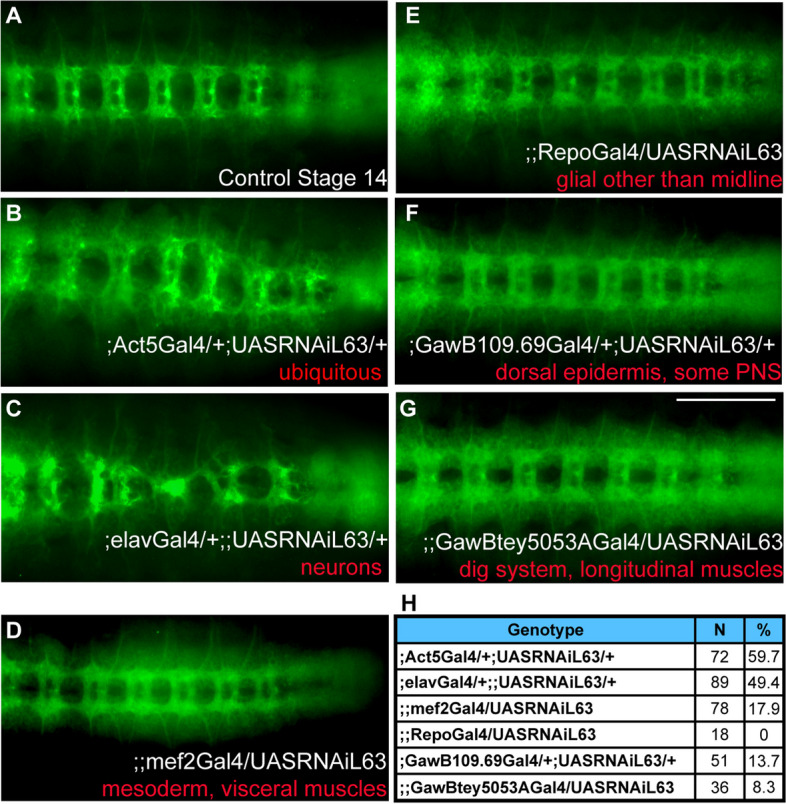
Background: PFTK1/Eip63E is a member of the cyclin-dependent kinases (CDKs) family and plays an important role in normal cell cycle progression. Eip63E expresses primarily in postnatal and adult nervous system in Drosophila melanogaster but its role in CNS development remains unknown. We sought to understand the function of Eip63E in the CNS by studying the fly ventral nerve cord during development.
Results: Our results demonstrate that Eip63E regulates axogenesis in neurons and its deficiency leads to neuronal defects. Functional interaction studies performed using the same system identify an interaction between Eip63E and the small GTPase Rho1. Furthermore, deficiency of Eip63E homolog in mice, PFTK1, in a newly generated PFTK1 knockout mice results in increased axonal outgrowth confirming that the developmental defects observed in the fly model are due to defects in axogenesis. Importantly, RhoA phosphorylation and activity are affected by PFTK1 in primary neuronal cultures. We report that GDP-bound inactive RhoA is a substrate of PFTK1 and PFTK1 phosphorylation is required for RhoA activity.
Conclusions: In conclusion, our work establishes an unreported neuronal role of PFTK1 in axon development mediated by phosphorylation and activation of GDP-bound RhoA. The results presented add to our understanding of the role of Cdks in the maintenance of RhoA-mediated axon growth and its impact on CNS development and axonal regeneration.
Keywords: Axogenesis; Cyclin-dependent kinases; Eip63E; Neuronal development; PFTAIRE; Rho.
© 2023. The Author(s).
Conflict of interest statement
PJ is a paid employee of Ionis Pharmaceuticals Inc. The other authors declare that they have no competing interests.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
