

Cryo-EM structure of the human cardiac myosin filament
- PMID: 37914935
- PMCID: PMC10846670
- DOI: 10.1038/s41586-023-06691-4
Cryo-EM structure of the human cardiac myosin filament
Abstract
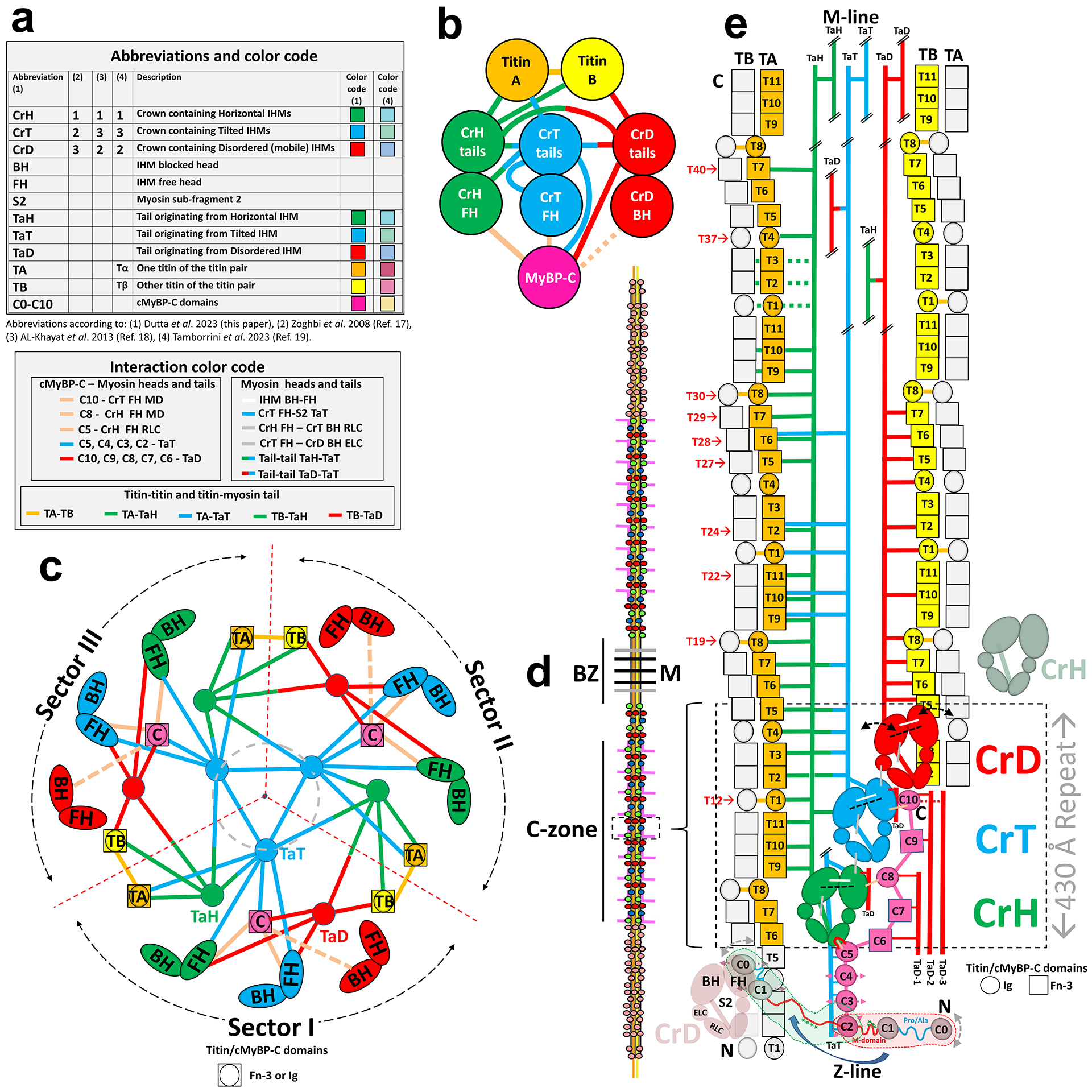
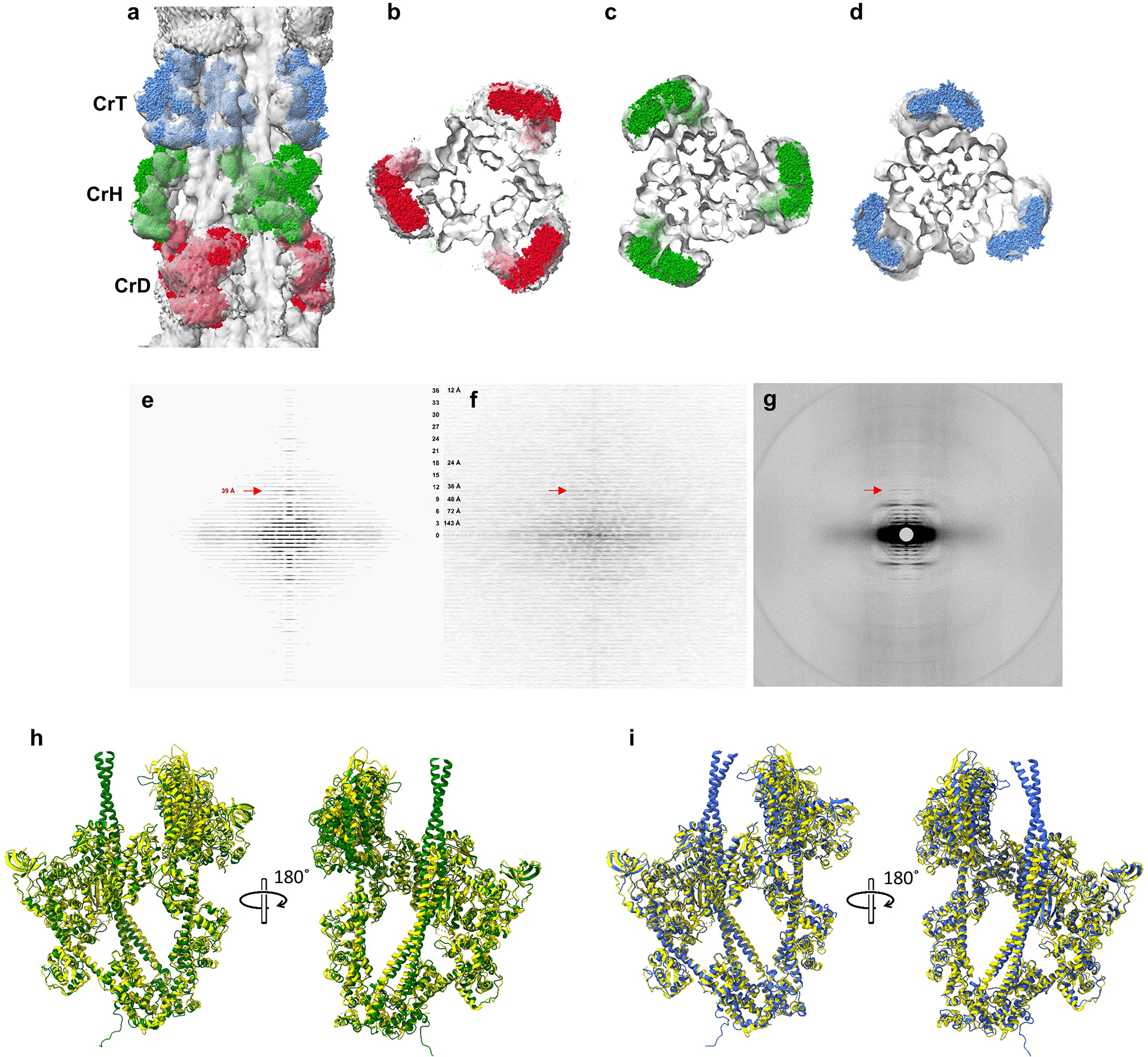
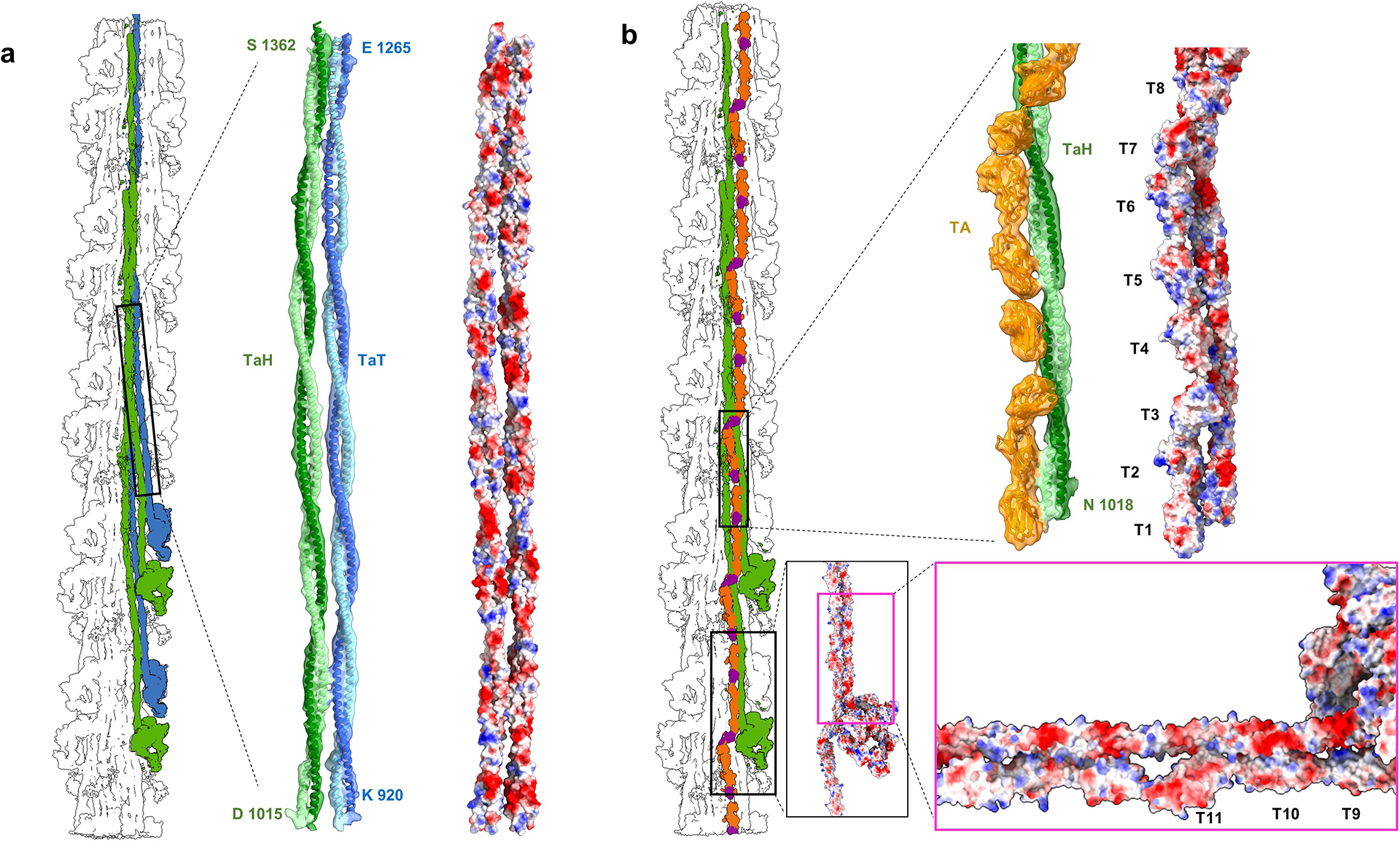
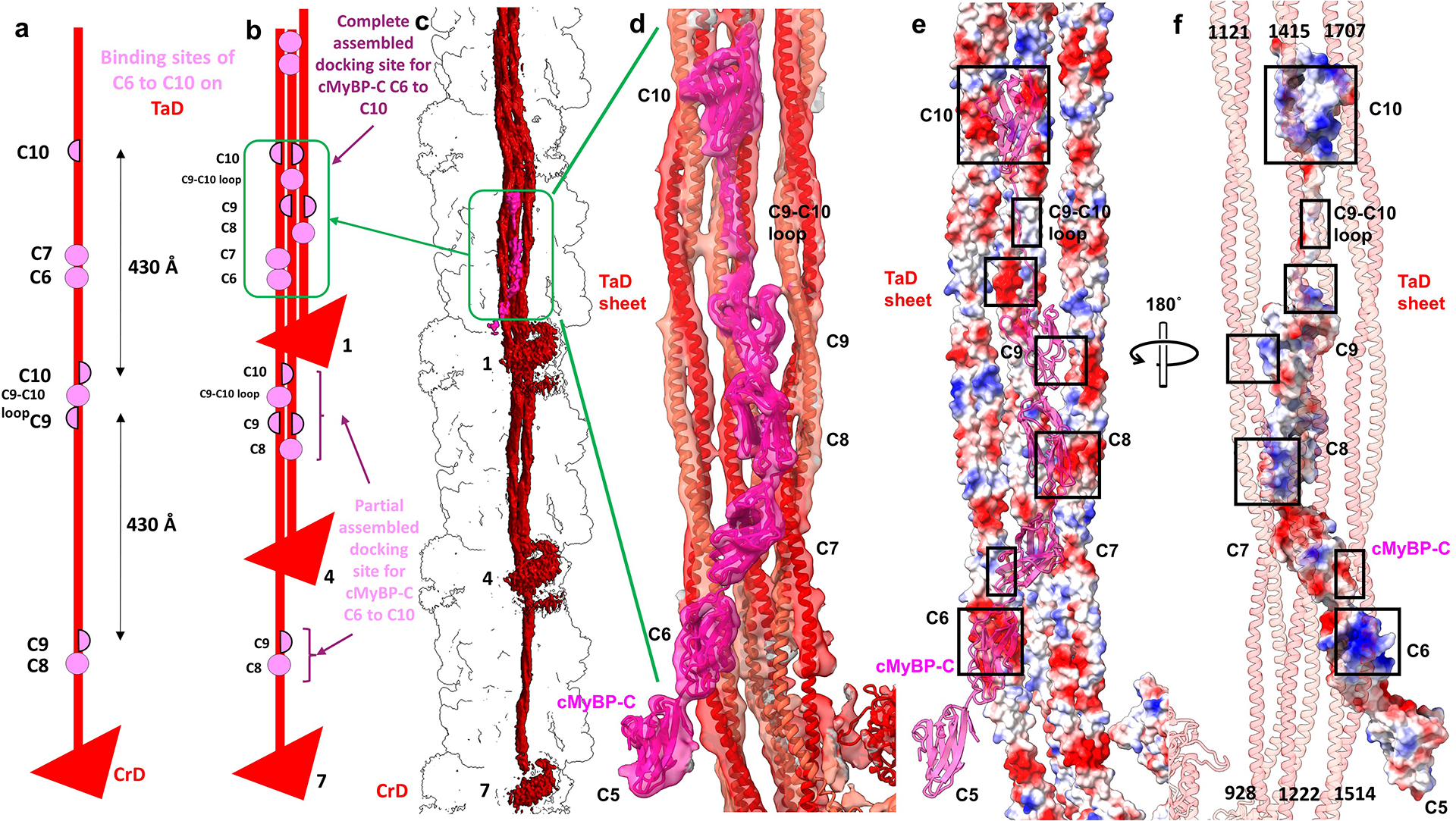
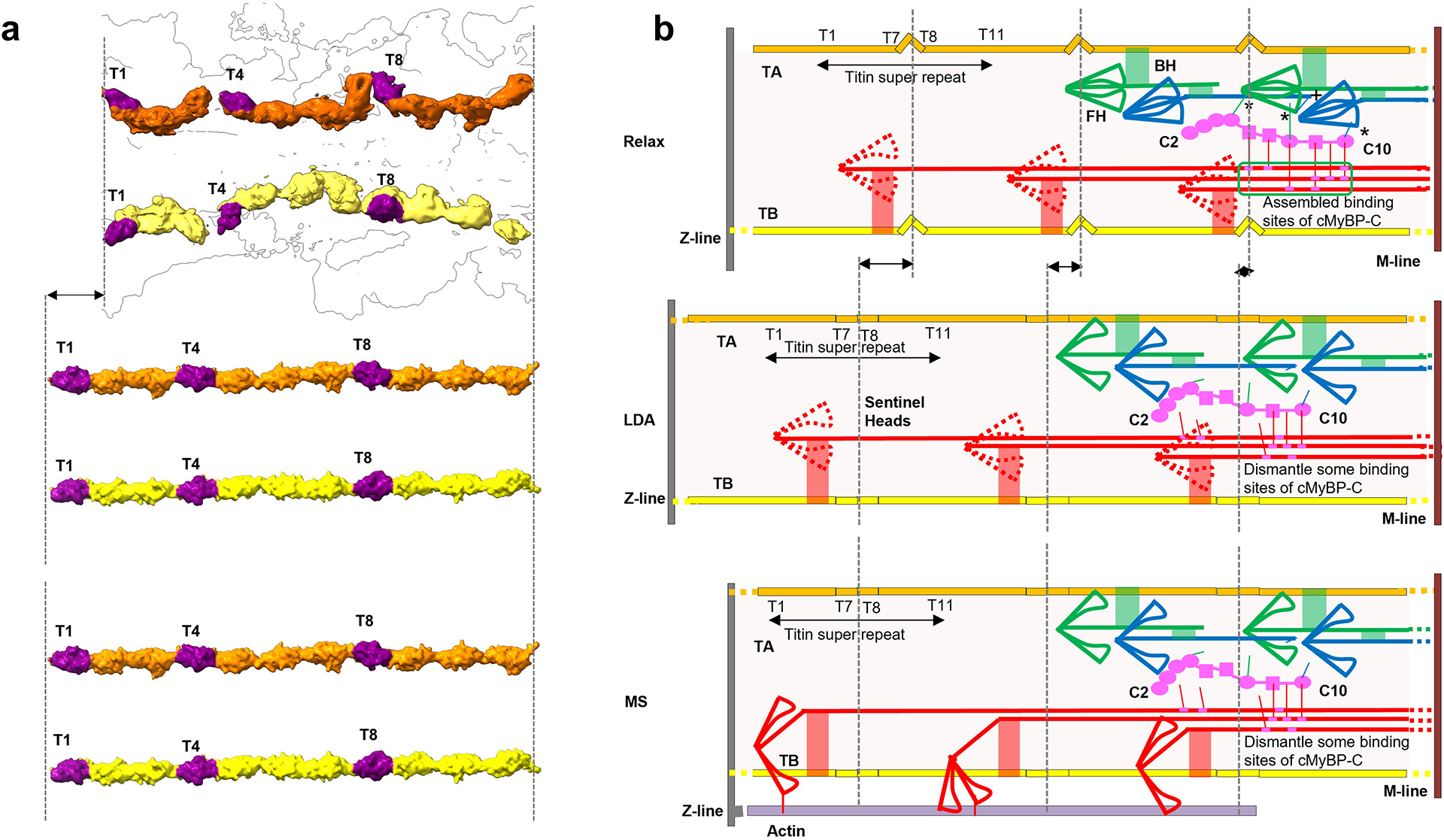
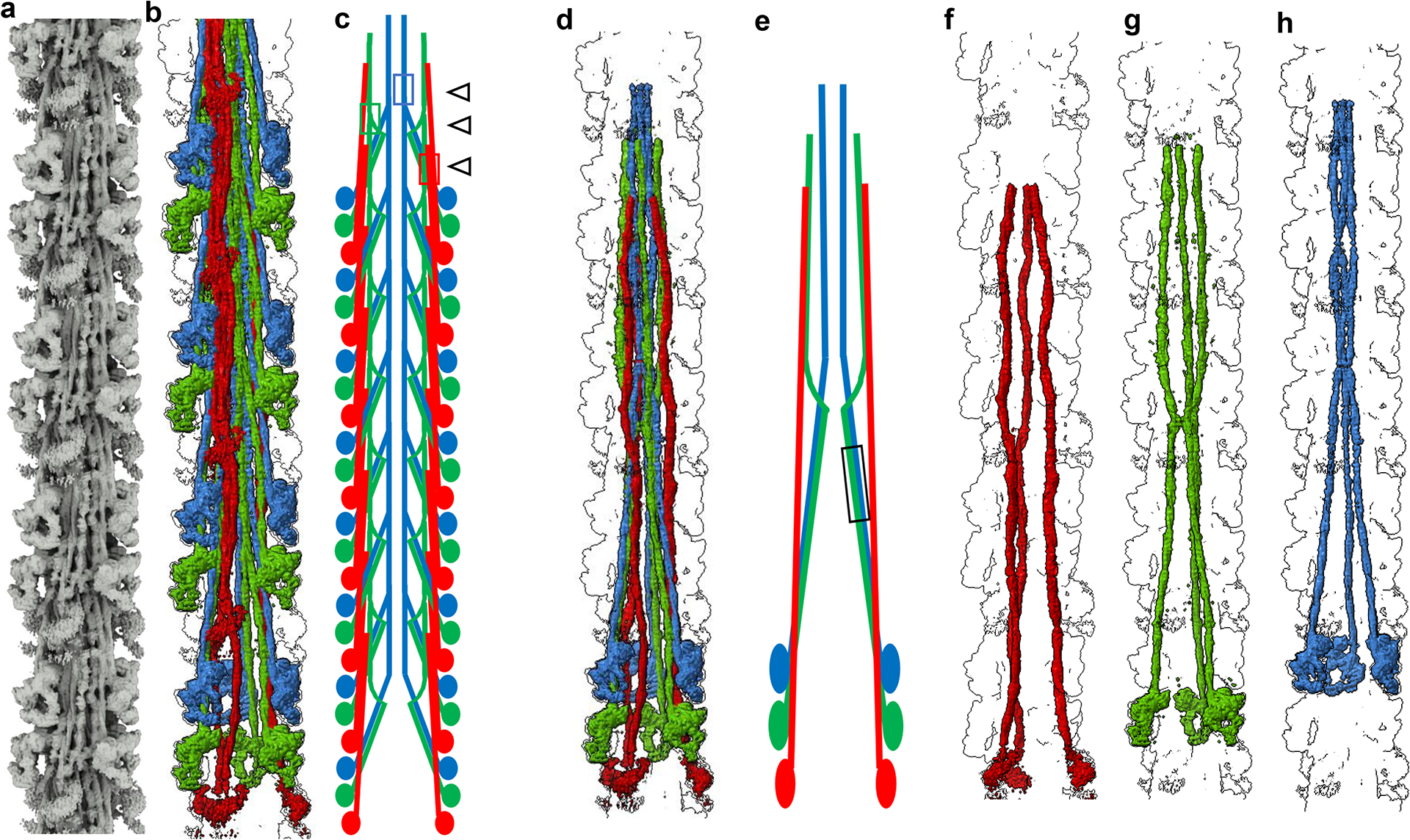
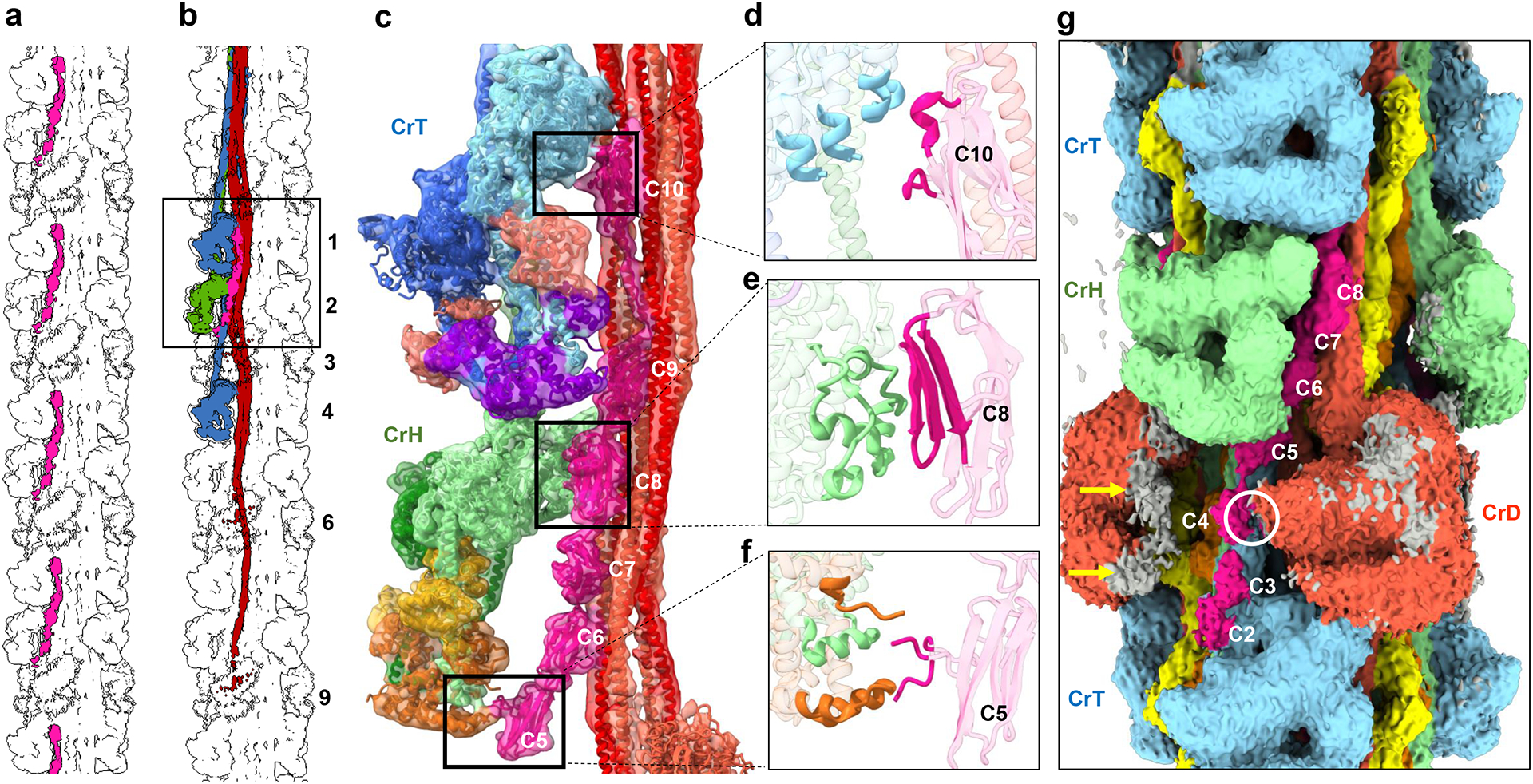
Pumping of the heart is powered by filaments of the motor protein myosin that pull on actin filaments to generate cardiac contraction. In addition to myosin, the filaments contain cardiac myosin-binding protein C (cMyBP-C), which modulates contractility in response to physiological stimuli, and titin, which functions as a scaffold for filament assembly1. Myosin, cMyBP-C and titin are all subject to mutation, which can lead to heart failure. Despite the central importance of cardiac myosin filaments to life, their molecular structure has remained a mystery for 60 years2. Here we solve the structure of the main (cMyBP-C-containing) region of the human cardiac filament using cryo-electron microscopy. The reconstruction reveals the architecture of titin and cMyBP-C and shows how myosin's motor domains (heads) form three different types of motif (providing functional flexibility), which interact with each other and with titin and cMyBP-C to dictate filament architecture and function. The packing of myosin tails in the filament backbone is also resolved. The structure suggests how cMyBP-C helps to generate the cardiac super-relaxed state3; how titin and cMyBP-C may contribute to length-dependent activation4; and how mutations in myosin and cMyBP-C might disturb interactions, causing disease5,6. The reconstruction resolves past uncertainties and integrates previous data on cardiac muscle structure and function. It provides a new paradigm for interpreting structural, physiological and clinical observations, and for the design of potential therapeutic drugs.
© 2023. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures









Update of
-
Cryo-EM structure of the human cardiac myosin filament.bioRxiv [Preprint]. 2023 Apr 12:2023.04.11.536274. doi: 10.1101/2023.04.11.536274. bioRxiv. 2023. Update in: Nature. 2023 Nov;623(7988):853-862. doi: 10.1038/s41586-023-06691-4. PMID: 37090534 Free PMC article. Updated. Preprint.
Comment in
-
Revealing the structure of the cardiac myosin filament.Nat Rev Cardiol. 2024 Jan;21(1):7. doi: 10.1038/s41569-023-00960-5. Nat Rev Cardiol. 2024. PMID: 37964080 No abstract available.
References
-
- Huxley HE Electron microscope studies on the structure of natural and synthetic protein filaments from striated muscle. J. Mol. Biol 7, 281–308 (1963). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
