

Single-haplotype comparative genomics provides insights into lineage-specific structural variation during cat evolution
- PMID: 37919451
- PMCID: PMC10845050
- DOI: 10.1038/s41588-023-01548-y
Single-haplotype comparative genomics provides insights into lineage-specific structural variation during cat evolution
Abstract
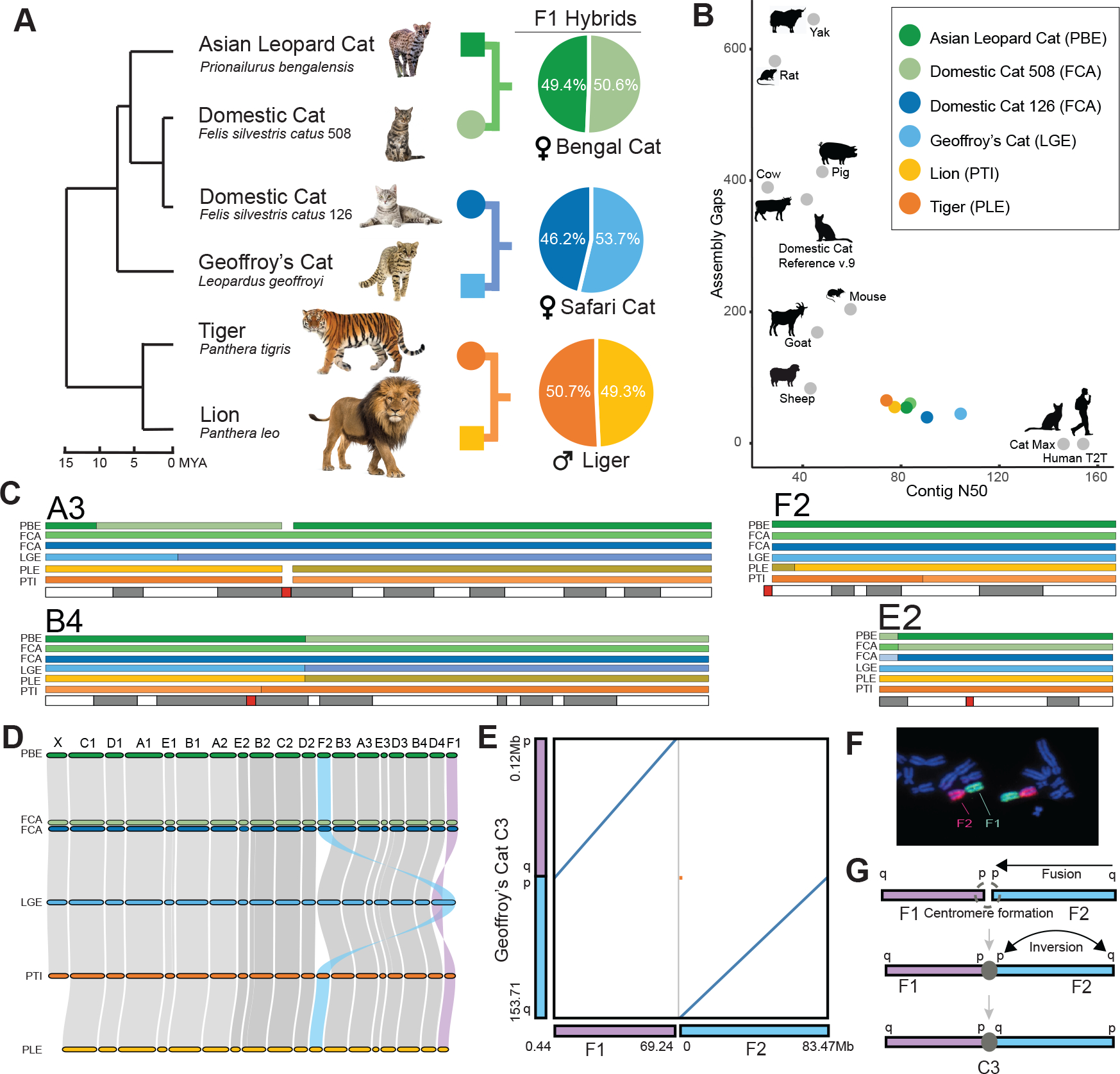
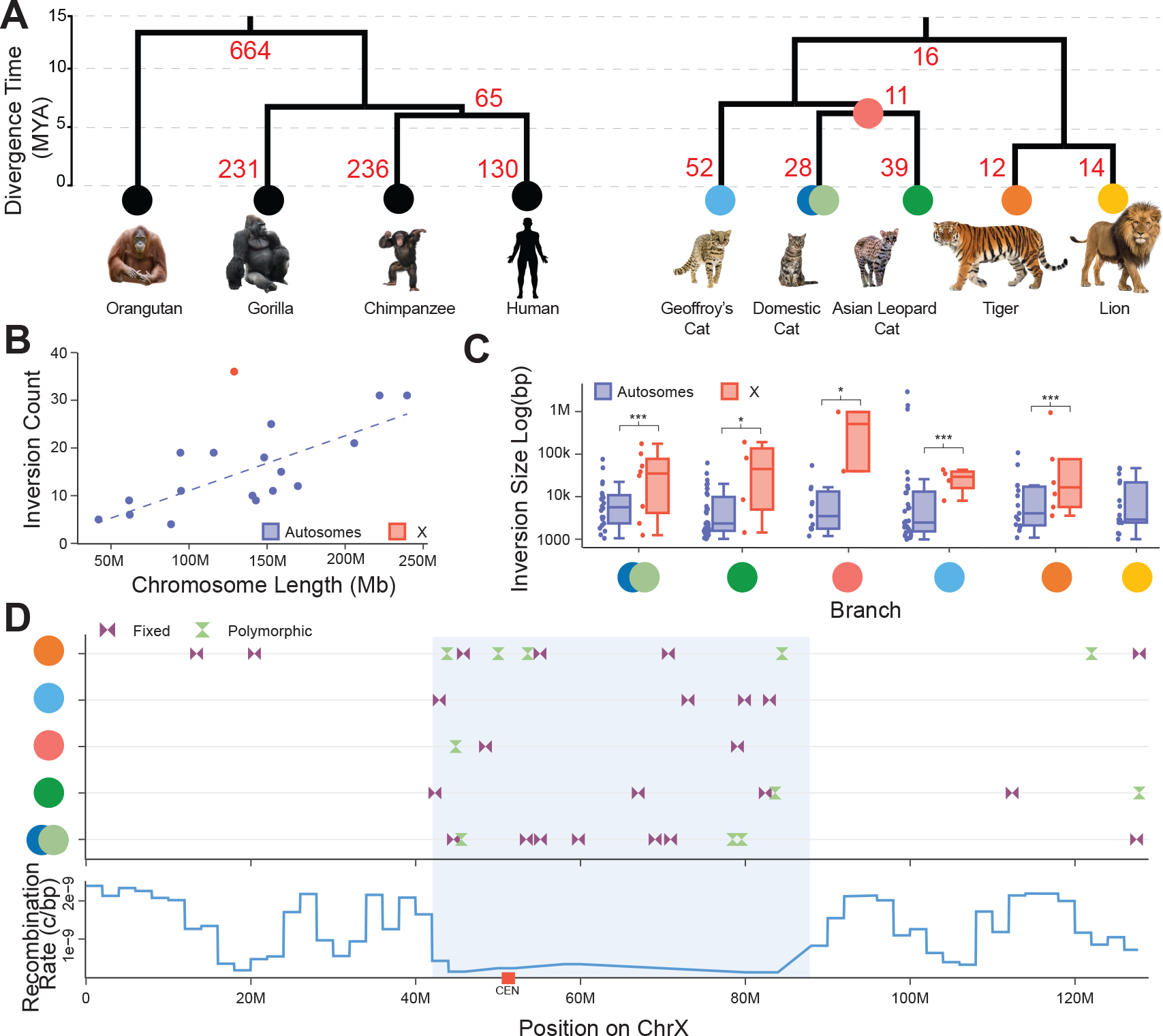
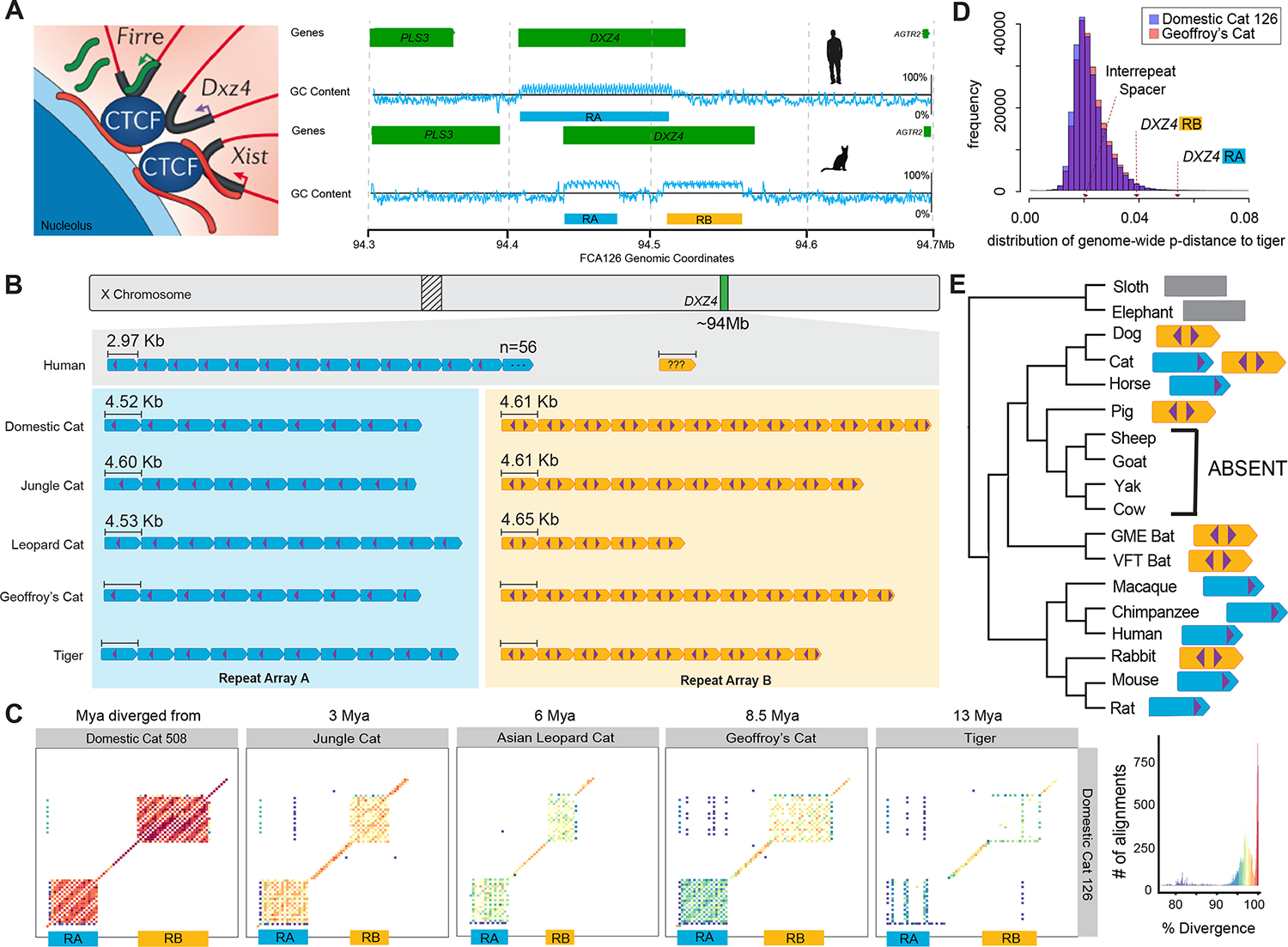
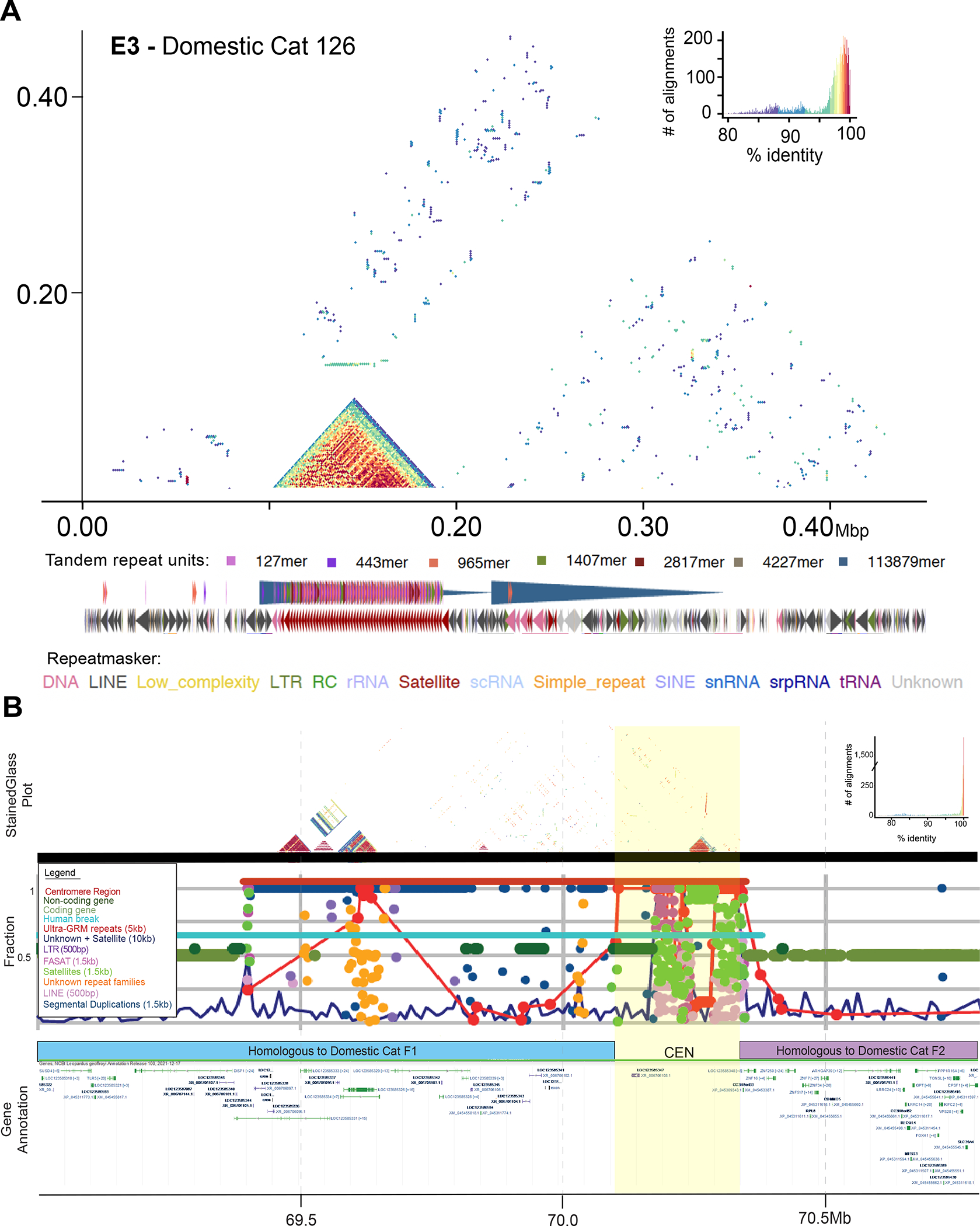
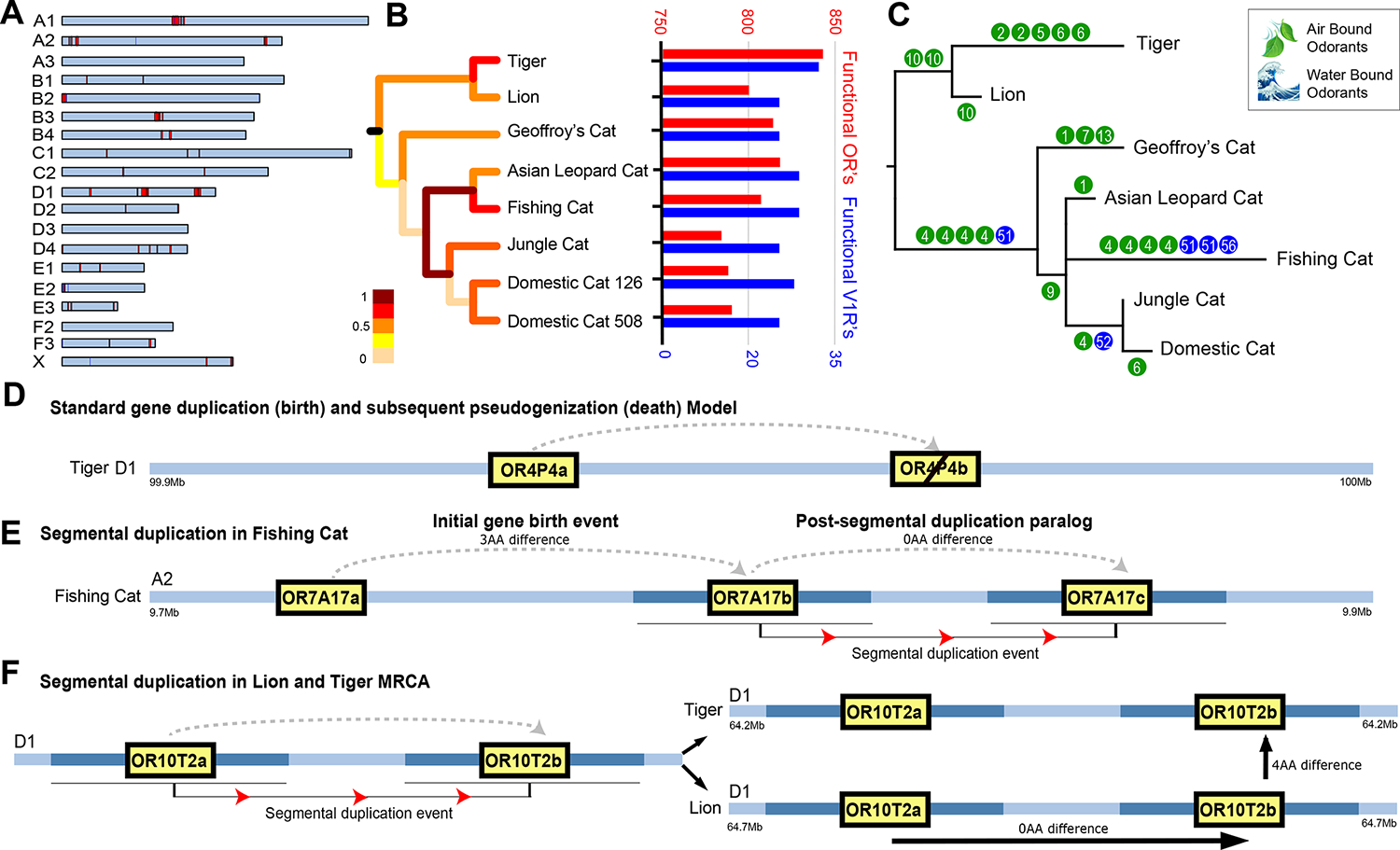
The role of structurally dynamic genomic regions in speciation is poorly understood due to challenges inherent in diploid genome assembly. Here we reconstructed the evolutionary dynamics of structural variation in five cat species by phasing the genomes of three interspecies F1 hybrids to generate near-gapless single-haplotype assemblies. We discerned that cat genomes have a paucity of segmental duplications relative to great apes, explaining their remarkable karyotypic stability. X chromosomes were hotspots of structural variation, including enrichment with inversions in a large recombination desert with characteristics of a supergene. The X-linked macrosatellite DXZ4 evolves more rapidly than 99.5% of the genome clarifying its role in felid hybrid incompatibility. Resolved sensory gene repertoires revealed functional copy number changes associated with ecomorphological adaptations, sociality and domestication. This study highlights the value of gapless genomes to reveal structural mechanisms underpinning karyotypic evolution, reproductive isolation and ecological niche adaptation.
© 2023. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Ethics declarations
Competing interests: The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
