

Development of functional resident macrophages in human pluripotent stem cell-derived colonic organoids and human fetal colon
- PMID: 37922878
- PMCID: PMC10913028
- DOI: 10.1016/j.stem.2023.10.002
Development of functional resident macrophages in human pluripotent stem cell-derived colonic organoids and human fetal colon
Abstract
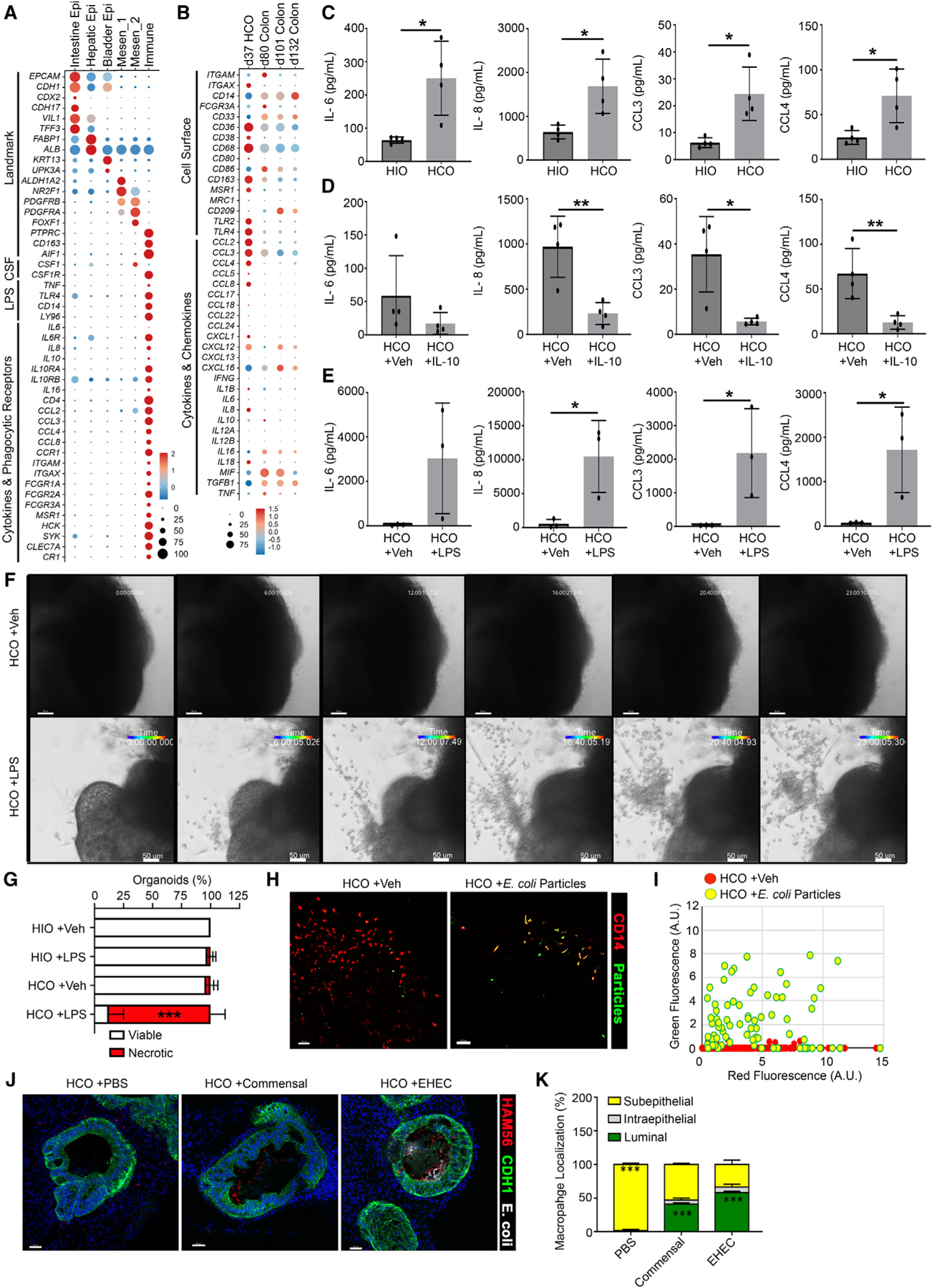
Most organs have tissue-resident immune cells. Human organoids lack these immune cells, which limits their utility in modeling many normal and disease processes. Here, we describe that pluripotent stem cell-derived human colonic organoids (HCOs) co-develop a diverse population of immune cells, including hemogenic endothelium (HE)-like cells and erythromyeloid progenitors that undergo stereotypical steps in differentiation, resulting in the generation of functional macrophages. HCO macrophages acquired a transcriptional signature resembling human fetal small and large intestine tissue-resident macrophages. HCO macrophages modulate cytokine secretion in response to pro- and anti-inflammatory signals and were able to phagocytose and mount a robust response to pathogenic bacteria. When transplanted into mice, HCO macrophages were maintained within the colonic organoid tissue, established a close association with the colonic epithelium, and were not displaced by the host bone-marrow-derived macrophages. These studies suggest that HE in HCOs gives rise to multipotent hematopoietic progenitors and functional tissue-resident macrophages.
Keywords: endothelial-to-hematopoietic transition; hemogenic endothelium; human colonic organoids; tissue-resident macrophages.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






References
Publication types
MeSH terms
Grants and funding
- UG3 DK119982/DK/NIDDK NIH HHS/United States
- T32 HL007574/HL/NHLBI NIH HHS/United States
- R01 DK137806/DK/NIDDK NIH HHS/United States
- P30 DK078392/DK/NIDDK NIH HHS/United States
- P20 GM130457/GM/NIGMS NIH HHS/United States
- UH3 DK119982/DK/NIDDK NIH HHS/United States
- U19 AI116491/AI/NIAID NIH HHS/United States
- T32 ES007250/ES/NIEHS NIH HHS/United States
- P30 DK123704/DK/NIDDK NIH HHS/United States
- U01 DK103117/DK/NIDDK NIH HHS/United States
- P01 HD093363/HD/NICHD NIH HHS/United States
- U18 EB021780/EB/NIBIB NIH HHS/United States
- RC2 DK120535/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
