

ATAC-clock: An aging clock based on chromatin accessibility
- PMID: 37924441
- PMCID: PMC10828344
- DOI: 10.1007/s11357-023-00986-0
ATAC-clock: An aging clock based on chromatin accessibility
Abstract
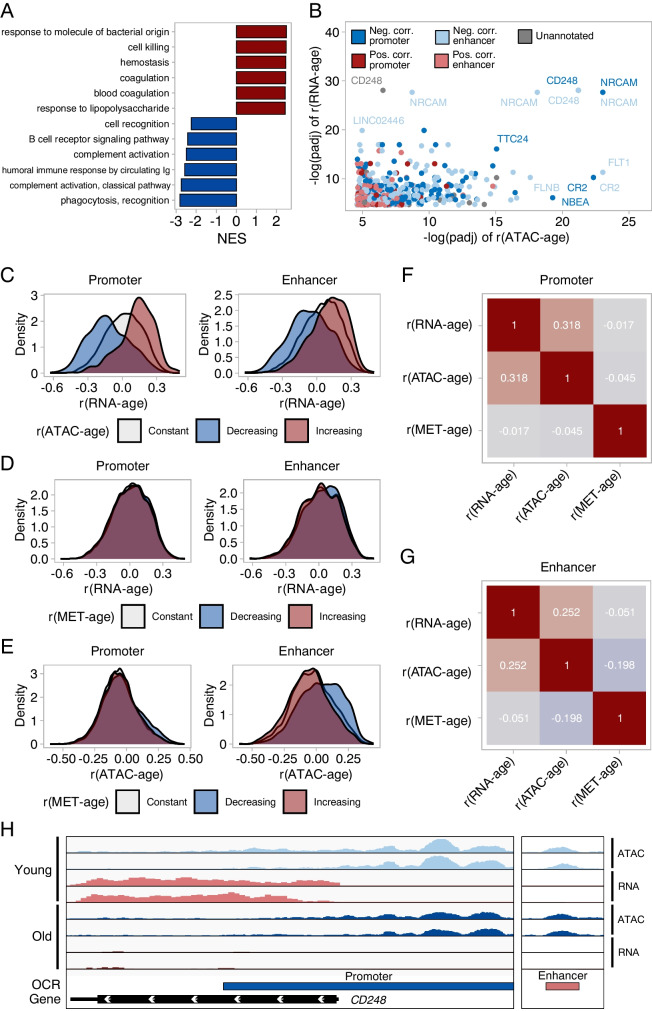
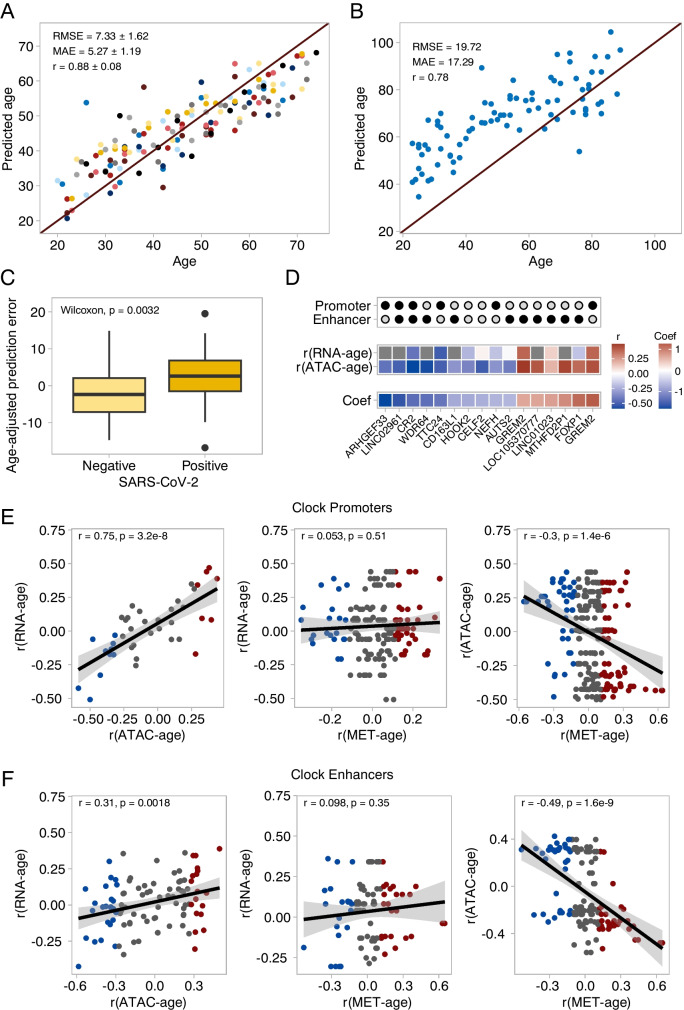
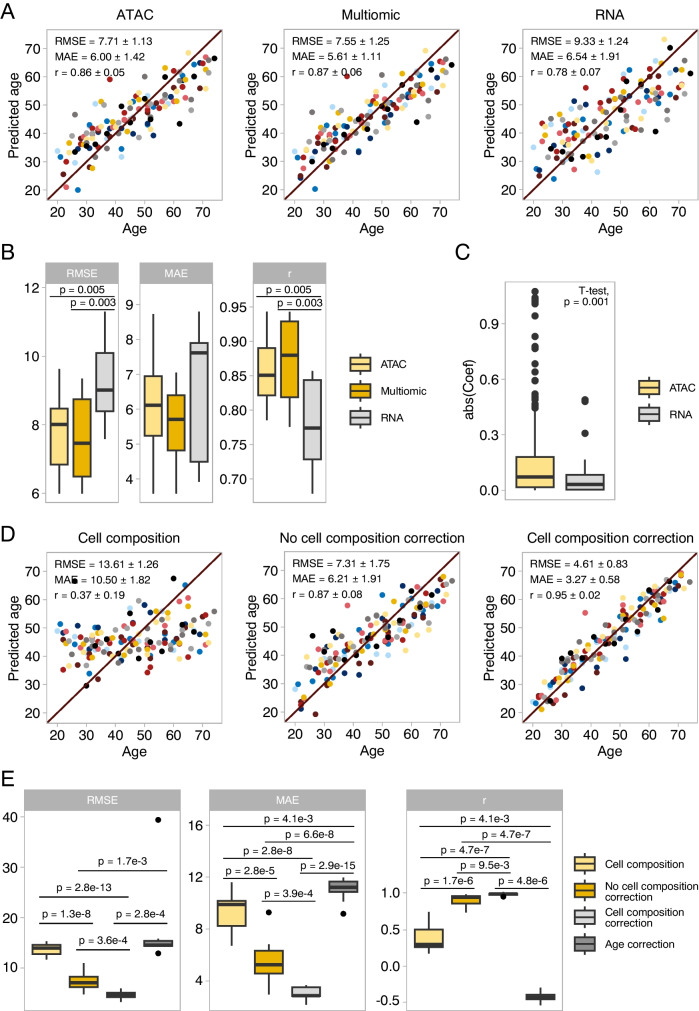
The establishment of aging clocks highlighted the strong link between changes in DNA methylation and aging. Yet, it is not known if other epigenetic features could be used to predict age accurately. Furthermore, previous studies have observed a lack of effect of age-related changes in DNA methylation on gene expression, putting the interpretability of DNA methylation-based aging clocks into question. In this study, we explore the use of chromatin accessibility to construct aging clocks. We collected blood from 159 human donors and generated chromatin accessibility, transcriptomic, and cell composition data. We investigated how chromatin accessibility changes during aging and constructed a novel aging clock with a median absolute error of 5.27 years. The changes in chromatin accessibility used by the clock were strongly related to transcriptomic alterations, aiding clock interpretation. We additionally show that our chromatin accessibility clock performs significantly better than a transcriptomic clock trained on matched samples. In conclusion, we demonstrate that the clock relies on cell-intrinsic chromatin accessibility alterations rather than changes in cell composition. Further, we present a new approach to construct epigenetic aging clocks based on chromatin accessibility, which bear a direct link to age-related transcriptional alterations, but which allow for more accurate age predictions than transcriptomic clocks.
Keywords: ATAC sequencing; Aging; Biomarker; Chromatin accessibility; Epigenetic clock.
© 2023. The Author(s).
Conflict of interest statement
K.P. and A.O. are co-founders and shareholders of EPITERNA SA (non-financial interests). A.O. is co-founder of Longevity Consultancy Group (non-financial interests). The rest of the authors have declared no financial or commercial conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
