

Differential responses to maternal diabetes in embryo and visceral yolk sac
- PMID: 37928898
- PMCID: PMC10620973
- DOI: 10.3389/fcell.2023.1273641
Differential responses to maternal diabetes in embryo and visceral yolk sac
Abstract
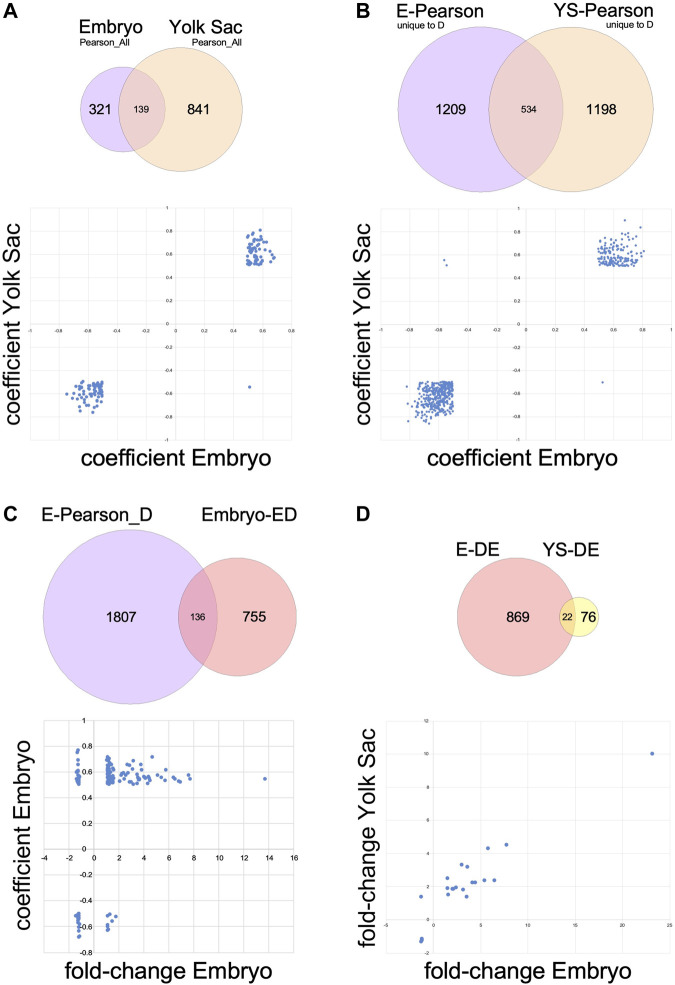
Introduction: Maternal diabetes during pregnancy is well known to be associated with a higher risk for structural birth defects in the offspring. Recent searches for underlying mechanisms have largely focused on aberrant processes in the embryo itself, although prior research in rodent models implicated dysfunction also of the visceral yolk sac. The objective of our research was to investigate both tissues within the conceptus simultaneously. Methods: We conducted unbiased transcriptome profiling by RNA sequencing on pairs of individual yolk sacs and their cognate embryos, using the non-obese diabetic (NOD) mouse model. The analysis was performed at gestational day 8.5 on morphologically normal specimen to circumvent confounding by defective development. Results: Even with large sample numbers (n = 33 in each group), we observed considerable variability of gene expression, primarily driven by exposure to maternal diabetes, and secondarily by developmental stage of the embryo. Only a moderate number of genes changed expression in the yolk sac, while in the embryo, the exposure distinctly influenced the relationship of gene expression levels to developmental progression, revealing a possible role for altered cell cycle regulation in the response. Also affected in embryos under diabetic conditions were genes involved in cholesterol biosynthesis and NAD metabolism pathways. Discussion: Exposure to maternal diabetes during gastrulation changes transcriptomic profiles in embryos to a substantially greater effect than in the corresponding yolk sacs, indicating that despite yolk sac being of embryonic origin, different mechanisms control transcriptional activity in these tissues. The effects of maternal diabetes on expression of many genes that are correlated with developmental progression (i.e. somite stage) highlight the importance of considering developmental maturity in the interpretation of transcriptomic data. Our analyses identified cholesterol biosynthesis and NAD metabolism as novel pathways not previously implicated in diabetic pregnancies. Both NAD and cholesterol availability affect a wide variety of cellular signaling processes, and can be modulated by diet, implying that prevention of adverse outcomes from diabetic pregnancies may require broad interventions, particularly in the early stages of pregnancy.
Keywords: RNA-seq; diabetic pregnancy; neural tube defect; non-obese diabetic mouse strain; somite number; somite stage; visceral yolk sac.
Copyright © 2023 Salbaum, Stone, Kruger and Kappen.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







References
-
- Cuny H., Rapadas M., Gereis J., Martin E., Kirk R. B., Shi H., et al. (2020). NAD deficiency due to environmental factors or gene-environment interactions causes congenital malformations and miscarriage in mice. Proc. Natl. Acad. Sci. U. S. A. 117, 3738–3747. 10.1073/pnas.1916588117 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases