

The DNA-binding induced (de)AMPylation activity of a Coxiella burnetii Fic enzyme targets Histone H3
- PMID: 37932372
- PMCID: PMC10628234
- DOI: 10.1038/s42003-023-05494-7
The DNA-binding induced (de)AMPylation activity of a Coxiella burnetii Fic enzyme targets Histone H3
Abstract
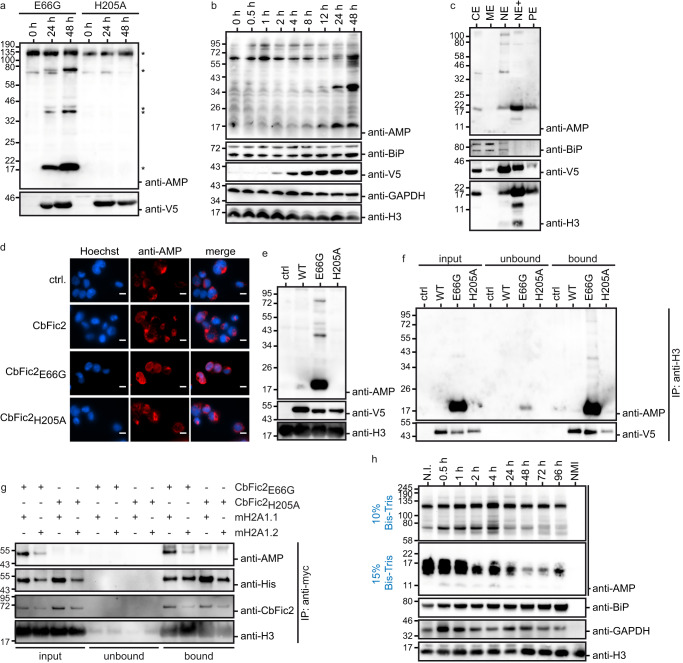
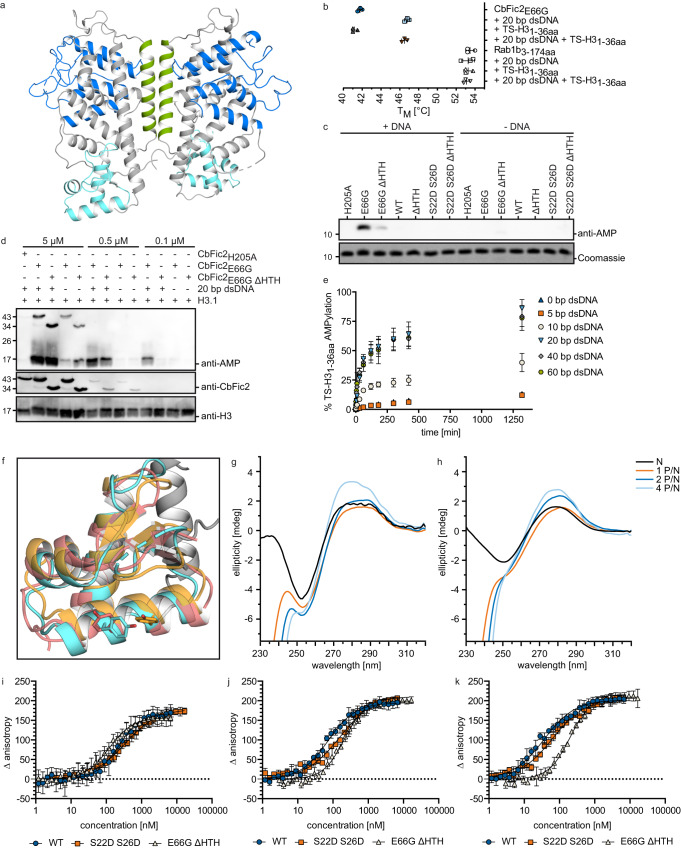
The intracellular bacterial pathogen Coxiella burnetii evades the host response by secreting effector proteins that aid in establishing a replication-friendly niche. Bacterial filamentation induced by cyclic AMP (Fic) enzymes can act as effectors by covalently modifying target proteins with the posttranslational AMPylation by transferring adenosine monophosphate (AMP) from adenosine triphosphate (ATP) to a hydroxyl-containing side chain. Here we identify the gene product of C. burnetii CBU_0822, termed C. burnetii Fic 2 (CbFic2), to AMPylate host cell histone H3 at serine 10 and serine 28. We show that CbFic2 acts as a bifunctional enzyme, both capable of AMPylation as well as deAMPylation, and is regulated by the binding of DNA via a C-terminal helix-turn-helix domain. We propose that CbFic2 performs AMPylation in its monomeric state, switching to a deAMPylating dimer upon DNA binding. This study unveils reversible histone modification by a specific enzyme of a pathogenic bacterium.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
