Contrasting diversity patterns of prokaryotes and protists over time and depth at the San-Pedro Ocean Time series
- PMID: 37938286
- PMCID: PMC9723720
- DOI: 10.1038/s43705-022-00121-8
Contrasting diversity patterns of prokaryotes and protists over time and depth at the San-Pedro Ocean Time series
Abstract
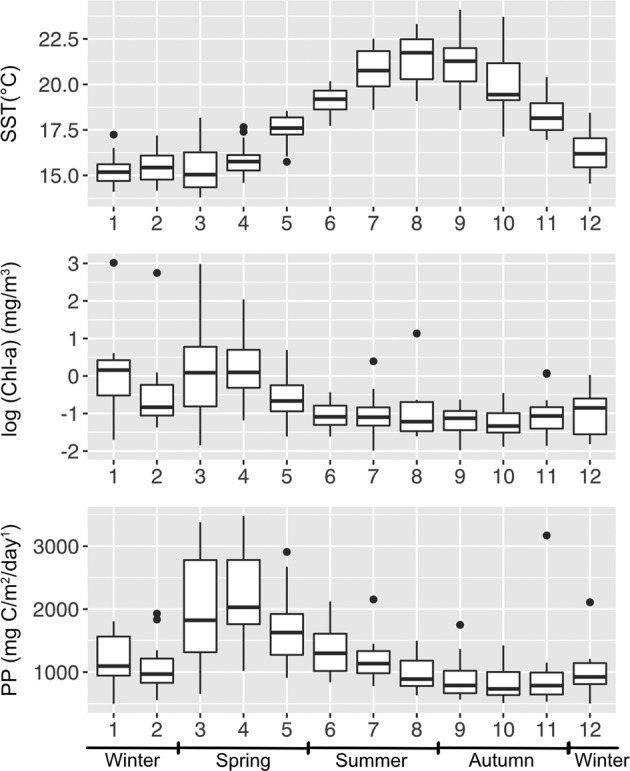
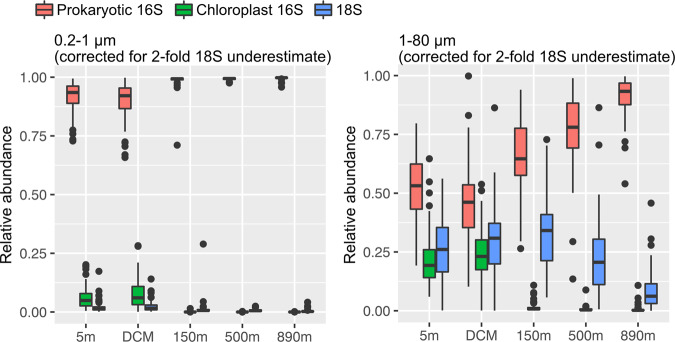
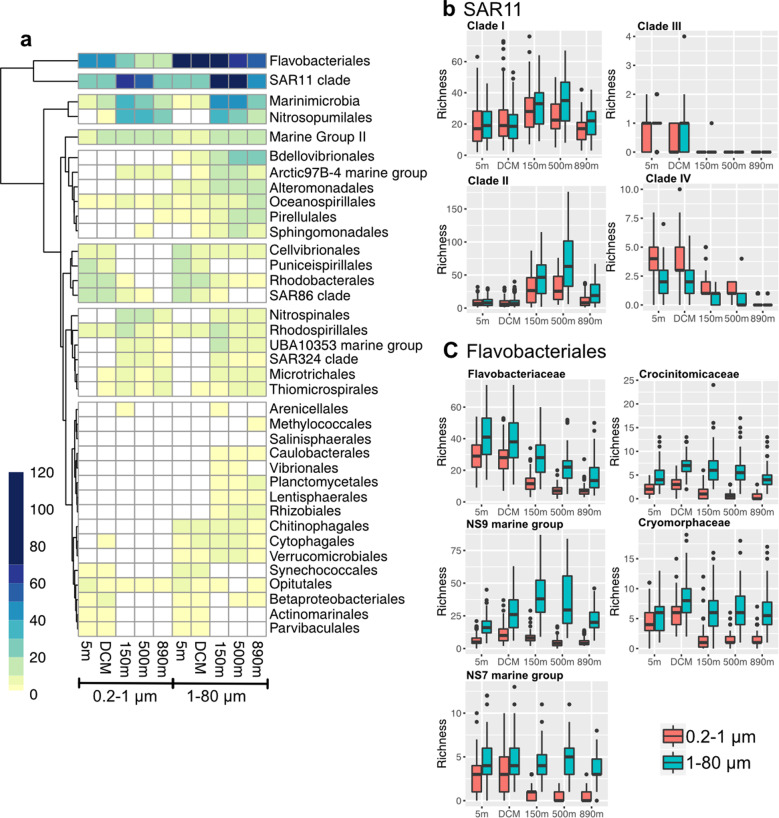
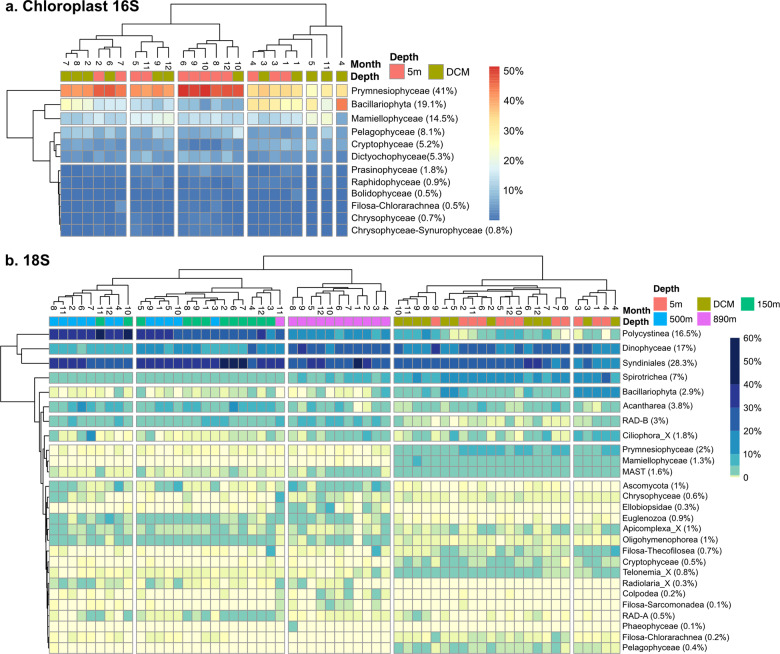
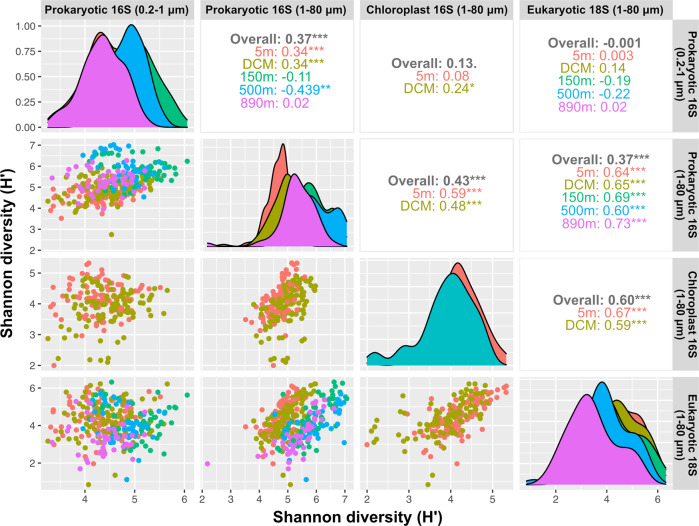
Community dynamics are central in microbial ecology, yet we lack studies comparing diversity patterns among marine protists and prokaryotes over depth and multiple years. Here, we characterized microbes at the San-Pedro Ocean Time series (2005-2018), using SSU rRNA gene sequencing from two size fractions (0.2-1 and 1-80 μm), with a universal primer set that amplifies from both prokaryotes and eukaryotes, allowing direct comparisons of diversity patterns in a single set of analyses. The 16S + 18S rRNA gene composition in the small size fraction was mostly prokaryotic (>92%) as expected, but the large size fraction unexpectedly contained 46-93% prokaryotic 16S rRNA genes. Prokaryotes and protists showed opposite vertical diversity patterns; prokaryotic diversity peaked at mid-depth, protistan diversity at the surface. Temporal beta-diversity patterns indicated prokaryote communities were much more stable than protists. Although the prokaryotic communities changed monthly, the average community stayed remarkably steady over 14 years, showing high resilience. Additionally, particle-associated prokaryotes were more diverse than smaller free-living ones, especially at deeper depths, contributed unexpectedly by abundant and diverse SAR11 clade II. Eukaryotic diversity was strongly correlated with the diversity of particle-associated prokaryotes but not free-living ones, reflecting that physical associations result in the strongest interactions, including symbioses, parasitism, and decomposer relationships.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures