

Plant carbonic anhydrase-like enzymes in neuroactive alkaloid biosynthesis
- PMID: 37938780
- PMCID: PMC10700139
- DOI: 10.1038/s41586-023-06716-y
Plant carbonic anhydrase-like enzymes in neuroactive alkaloid biosynthesis
Abstract
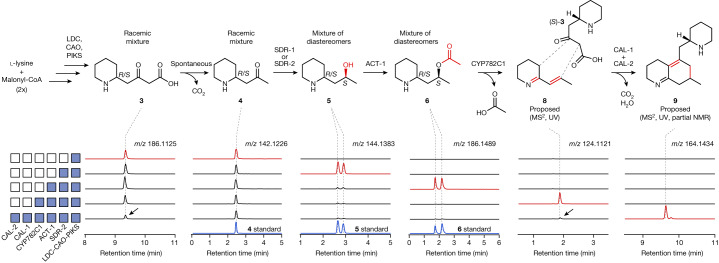
Plants synthesize numerous alkaloids that mimic animal neurotransmitters1. The diversity of alkaloid structures is achieved through the generation and tailoring of unique carbon scaffolds2,3, yet many neuroactive alkaloids belong to a scaffold class for which no biosynthetic route or enzyme catalyst is known. By studying highly coordinated, tissue-specific gene expression in plants that produce neuroactive Lycopodium alkaloids4, we identified an unexpected enzyme class for alkaloid biosynthesis: neofunctionalized α-carbonic anhydrases (CAHs). We show that three CAH-like (CAL) proteins are required in the biosynthetic route to a key precursor of the Lycopodium alkaloids by catalysing a stereospecific Mannich-like condensation and subsequent bicyclic scaffold generation. Also, we describe a series of scaffold tailoring steps that generate the optimized acetylcholinesterase inhibition activity of huperzine A5. Our findings suggest a broader involvement of CAH-like enzymes in specialized metabolism and demonstrate how successive scaffold tailoring can drive potency against a neurological protein target.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures














Comment in
-
Plant gene co-expression defines the biosynthetic pathway of neuroactive alkaloids.Mol Plant. 2024 Mar 4;17(3):372-374. doi: 10.1016/j.molp.2024.02.002. Epub 2024 Feb 6. Mol Plant. 2024. PMID: 38321739 No abstract available.
References
-
- Wink, M. in Modern Alkaloids: Structure, Isolation, Synthesis and Biology (eds Fattorusso, E. & Taglialatela‐Scafati, O.) 1–24 (Wiley, 2007).
-
- Liu J-S, et al. The structures of huperzine A and B, two new alkaloids exhibiting marked anticholinesterase activity. Can. J. Chem. 1986;64:837–839. doi: 10.1139/v86-137. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
