

Inhibition of striatal dopamine release by the L-type calcium channel inhibitor isradipine co-varies with risk factors for Parkinson's
- PMID: 37941514
- PMCID: PMC11426196
- DOI: 10.1111/ejn.16180
Inhibition of striatal dopamine release by the L-type calcium channel inhibitor isradipine co-varies with risk factors for Parkinson's
Abstract
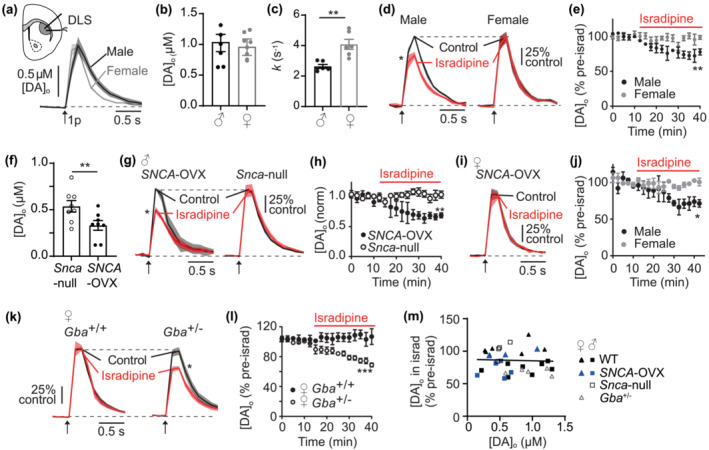
Ca2+ entry into nigrostriatal dopamine (DA) neurons and axons via L-type voltage-gated Ca2+ channels (LTCCs) contributes, respectively, to pacemaker activity and DA release and has long been thought to contribute to vulnerability to degeneration in Parkinson's disease. LTCC function is greater in DA axons and neurons from substantia nigra pars compacta than from ventral tegmental area, but this is not explained by channel expression level. We tested the hypothesis that LTCC control of DA release is governed rather by local mechanisms, focussing on candidate biological factors known to operate differently between types of DA neurons and/or be associated with their differing vulnerability to parkinsonism, including biological sex, α-synuclein, DA transporters (DATs) and calbindin-D28k (Calb1). We detected evoked DA release ex vivo in mouse striatal slices using fast-scan cyclic voltammetry and assessed LTCC support of DA release by detecting the inhibition of DA release by the LTCC inhibitors isradipine or CP8. Using genetic knockouts or pharmacological manipulations, we identified that striatal LTCC support of DA release depended on multiple intersecting factors, in a regionally and sexually divergent manner. LTCC function was promoted by factors associated with Parkinsonian risk, including male sex, α-synuclein, DAT and a dorsolateral co-ordinate, but limited by factors associated with protection, that is, female sex, glucocerebrosidase activity, Calb1 and ventromedial co-ordinate. Together, these data show that LTCC function in DA axons and isradipine effect are locally governed and suggest they vary in a manner that in turn might impact on, or reflect, the cellular stress that leads to parkinsonian degeneration.
Keywords: Calb1; L‐type calcium channel; Parkinson's disease; dopamine D2 receptor; dopamine release; dopamine transporter; sex differences; striatum; synuclein.
© 2023 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Conflict of interest statement
The authors have no conflicts of interest to disclose.
Figures




References
-
- Airaksinen, M. S. , Thoenen, H. , & Meyer, M. (1997). Vulnerability of midbrain dopaminergic neurons in Calbindin‐D 28k ‐deficient mice: Lack of evidence for a neuroprotective role of endogenous Calbindin in MPTPtreated and weaver mice. The European Journal of Neuroscience, 9, 120–127. 10.1111/j.1460-9568.1997.tb01360.x - DOI - PubMed
-
- Alix, J. J. P. , Dolphin, A. C. , & Fern, R. (2008). Vesicular apparatus, including functional calcium channels, are present in developing rodent optic nerve axons and are required for normal node of Ranvier formation. The Journal of Physiology, 586, 4069–4089. 10.1113/jphysiol.2008.155077 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
