

Nuclear transport proteins: structure, function, and disease relevance
- PMID: 37945593
- PMCID: PMC10636164
- DOI: 10.1038/s41392-023-01649-4
Nuclear transport proteins: structure, function, and disease relevance
Abstract
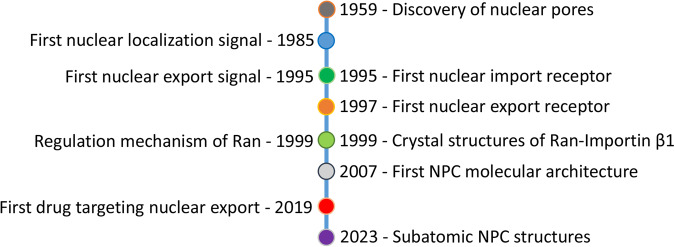
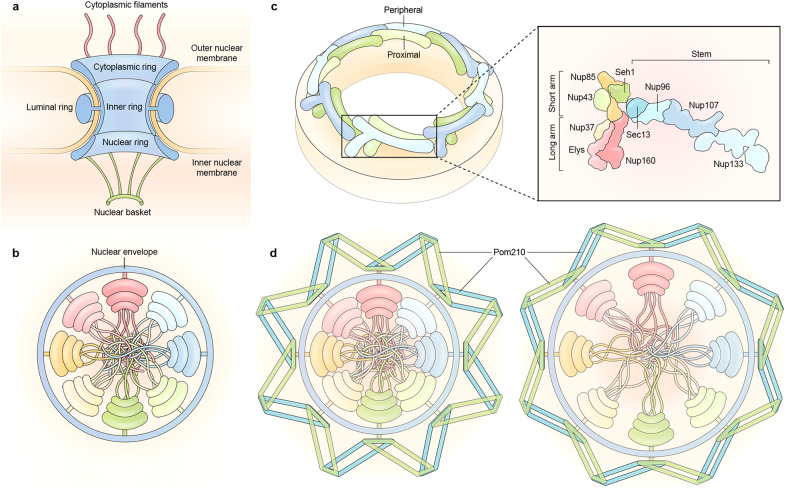
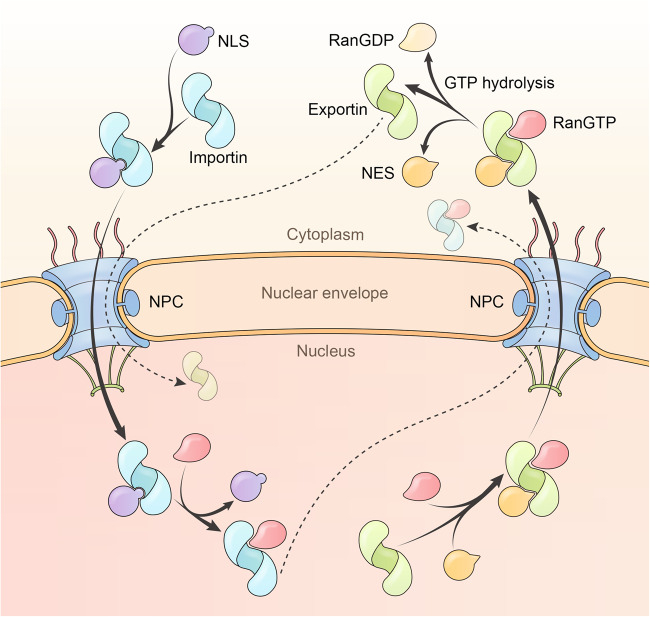
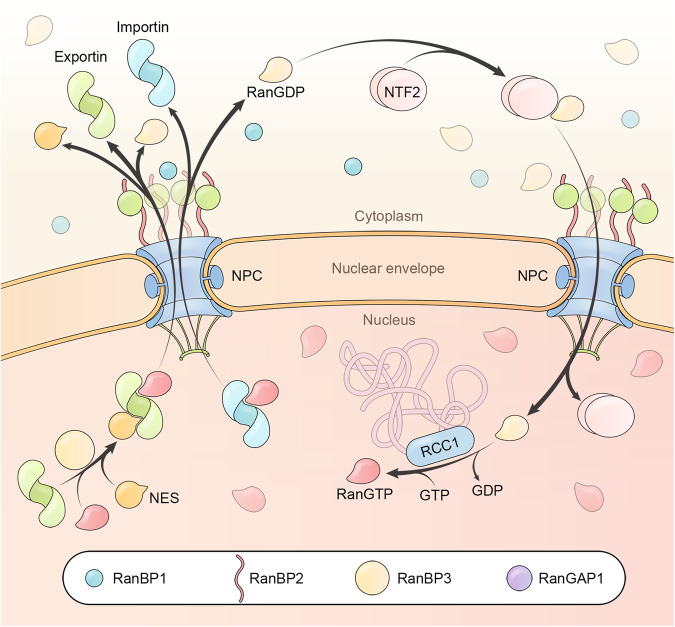
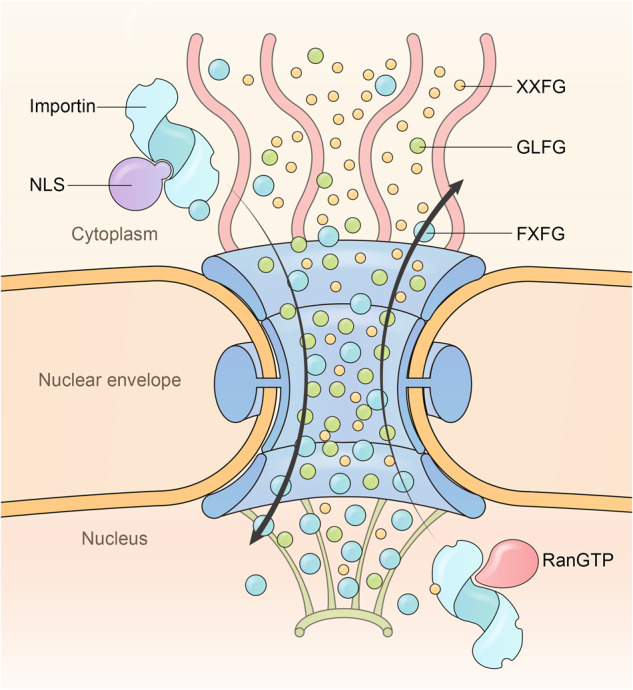
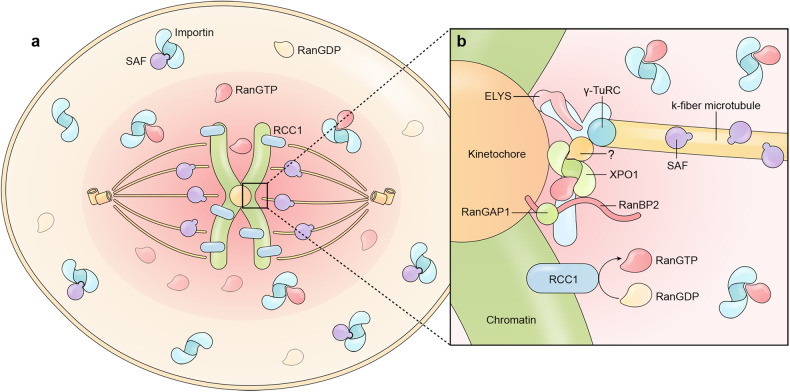
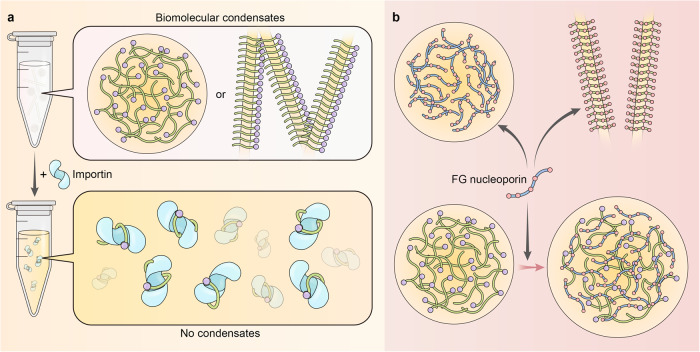
Proper subcellular localization is crucial for the functioning of biomacromolecules, including proteins and RNAs. Nuclear transport is a fundamental cellular process that regulates the localization of many macromolecules within the nuclear or cytoplasmic compartments. In humans, approximately 60 proteins are involved in nuclear transport, including nucleoporins that form membrane-embedded nuclear pore complexes, karyopherins that transport cargoes through these complexes, and Ran system proteins that ensure directed and rapid transport. Many of these nuclear transport proteins play additional and essential roles in mitosis, biomolecular condensation, and gene transcription. Dysregulation of nuclear transport is linked to major human diseases such as cancer, neurodegenerative diseases, and viral infections. Selinexor (KPT-330), an inhibitor targeting the nuclear export factor XPO1 (also known as CRM1), was approved in 2019 to treat two types of blood cancers, and dozens of clinical trials of are ongoing. This review summarizes approximately three decades of research data in this field but focuses on the structure and function of individual nuclear transport proteins from recent studies, providing a cutting-edge and holistic view on the role of nuclear transport proteins in health and disease. In-depth knowledge of this rapidly evolving field has the potential to bring new insights into fundamental biology, pathogenic mechanisms, and therapeutic approaches.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Beck, M. & Hurt, E. The nuclear pore complex: understanding its function through structural insight. Nat. Rev. Mol. Cell Biol.18, 73–89 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
