

Irf7 regulates the expression of Srg3 and ferroptosis axis aggravated sepsis-induced acute lung injury
- PMID: 37946128
- PMCID: PMC10634032
- DOI: 10.1186/s11658-023-00495-0
Irf7 regulates the expression of Srg3 and ferroptosis axis aggravated sepsis-induced acute lung injury
Abstract
Objective: To investigate the mechanism of action of Srg3 in acute lung injury caused by sepsis.
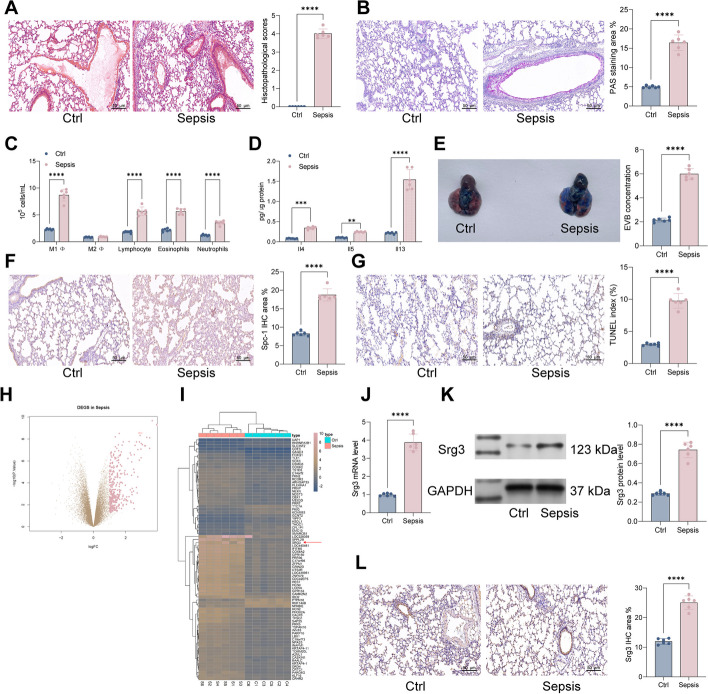
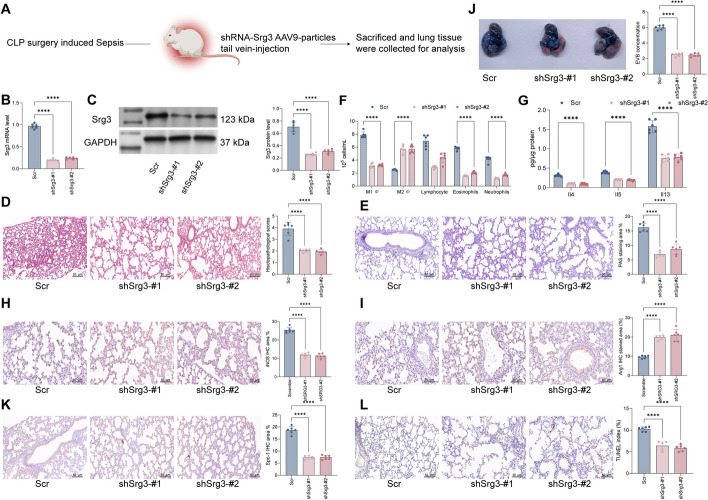
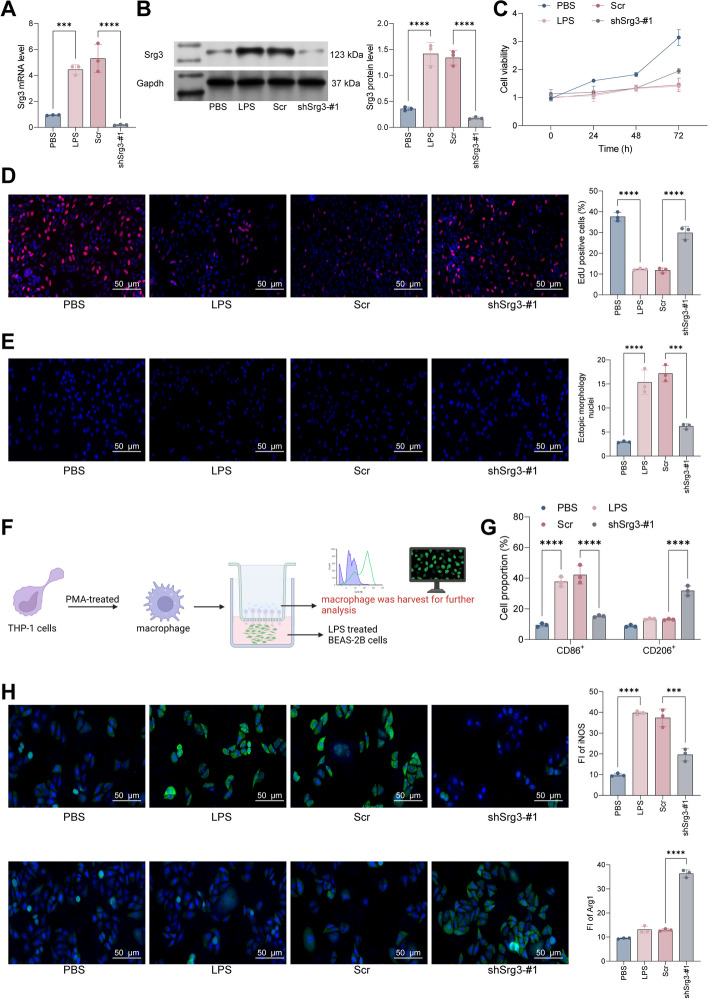
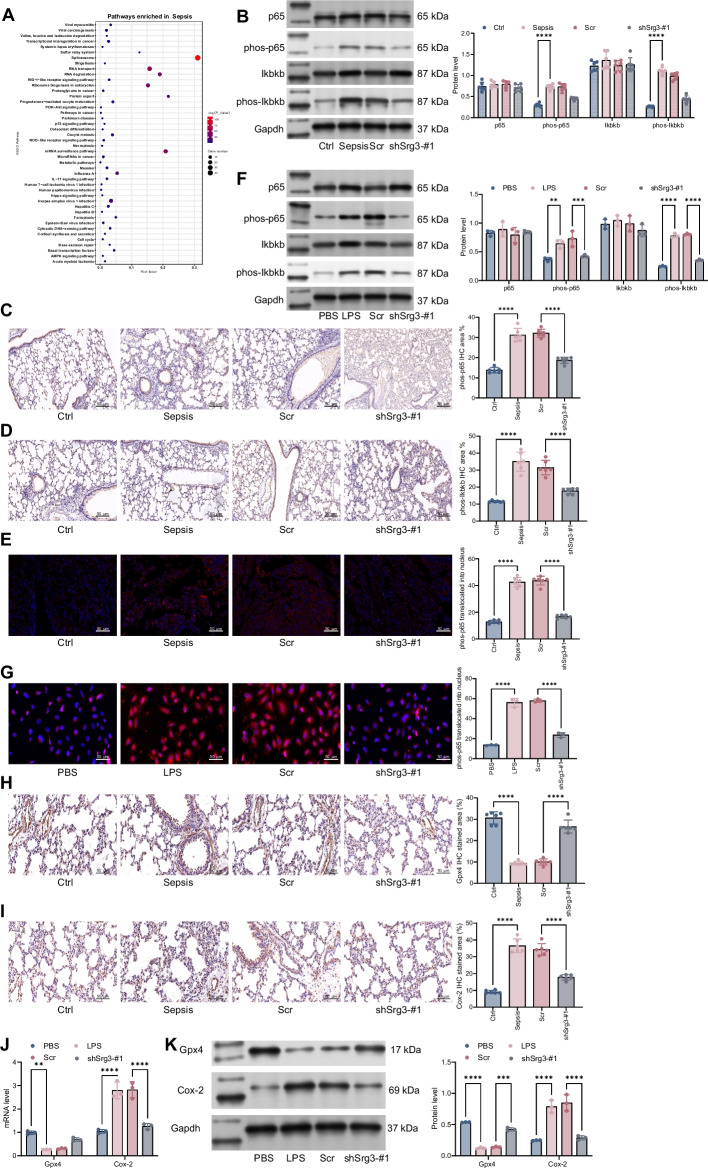
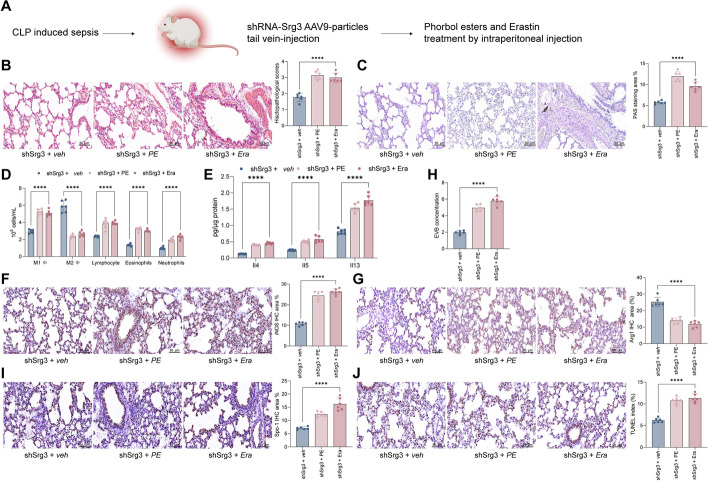
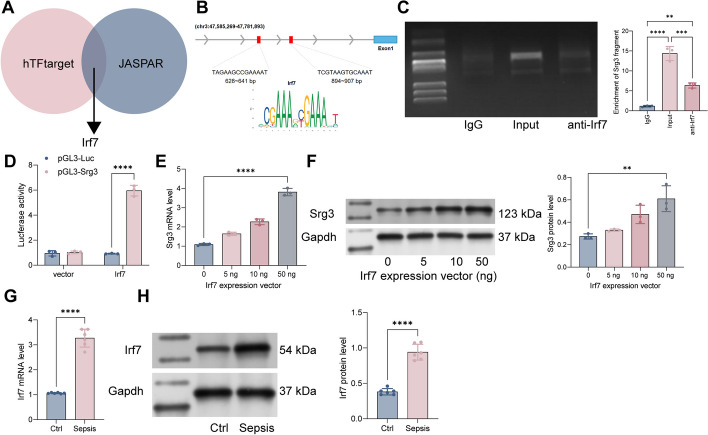
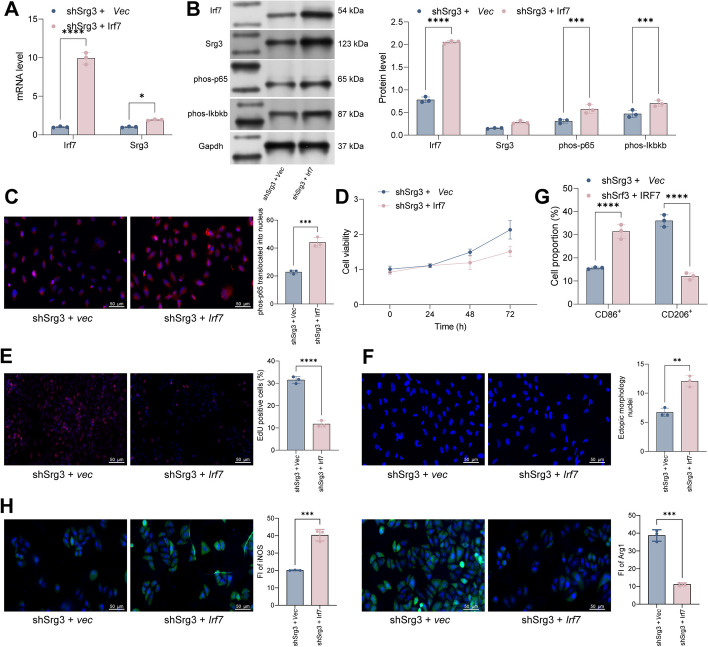
Methods: First, a sepsis-induced acute lung injury rat model was established using cecal ligation and puncture (CLP). RNA sequencing (RNA-seq) was used to screen for highly expressed genes in sepsis-induced acute lung injury (ALI), and the results showed that Srg3 was significantly upregulated. Then, SWI3-related gene 3 (Srg3) was knocked down using AAV9 vector in vivo, and changes in ALI symptoms in rats were analyzed. In vitro experiments were conducted by establishing a cell model using lipopolysaccharide (LPS)-induced BEAS-2B cells and coculturing them with phorbol 12-myristate 13-acetate (PMA)-treated THP-1 cells to analyze macrophage polarization. Next, downstream signaling pathways regulated by Srg3 and transcription factors involved in regulating Srg3 expression were analyzed using the KEGG database. Finally, gain-of-loss functional validation experiments were performed to analyze the role of downstream signaling pathways regulated by Srg3 and transcription factors involved in regulating Srg3 expression in sepsis-induced acute lung injury.
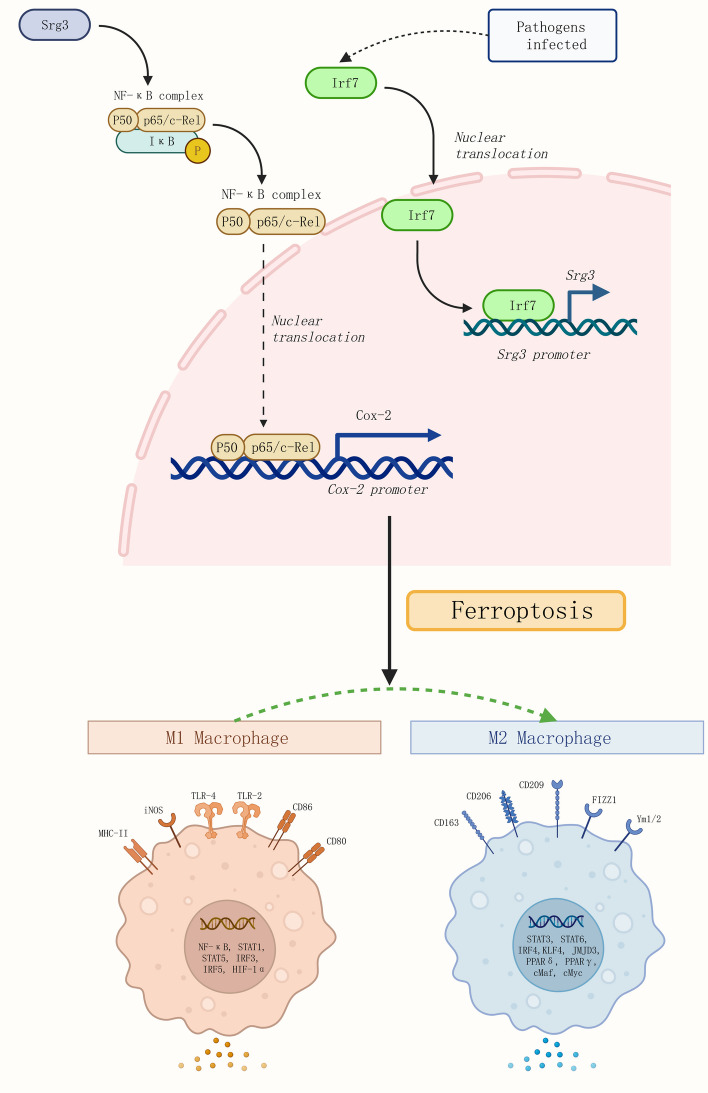
Results: Srg3 was significantly upregulated in sepsis-induced acute lung injury, and knocking down Srg3 significantly improved the symptoms of ALI in rats. Furthermore, in vitro experiments showed that knocking down Srg3 significantly weakened the inhibitory effect of LPS on the viability of BEAS-2B cells and promoted alternative activation phenotype (M2) macrophage polarization. Subsequent experiments showed that Srg3 can regulate the activation of the NF-κB signaling pathway and promote ferroptosis. Specific activation of the NF-κB signaling pathway or ferroptosis significantly weakened the effect of Srg3 knockdown. It was then found that Srg3 can be transcriptionally activated by interferon regulatory factor 7 (Irf7), and specific inhibition of Irf7 significantly improved the symptoms of ALI.
Conclusions: Irf7 transcriptionally activates the expression of Srg3, which can promote ferroptosis and activate classical activation phenotype (M1) macrophage polarization by regulating the NF-κB signaling pathway, thereby exacerbating the symptoms of septic lung injury.
Keywords: Ferroptosis; Irf7; NF-κB signaling pathway; Sepsis-induced acute lung injury; Srg3.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Fleischmann-Struzek C, Mellhammar L, Rose N, Cassini A, Rudd KE, Schlattmann P, et al. Incidence and mortality of hospital- and ICU-treated sepsis: Results from an updated and expanded systematic review and meta-analysis. Intensive Care Med. 2020;46(8):1552–1562. doi: 10.1007/s00134-020-06151-x. - DOI - PMC - PubMed
-
- Gotts JE, Matthay MA. Sepsis: Pathophysiology and clinical management. BMJ (Clinical research ed) 2016;353:i1585. - PubMed
-
- Wang W. The SWI/SNF family of ATP-dependent chromatin remodelers: Similar mechanisms for diverse functions. Curr Top Microbiol Immunol. 2003;274:143–169. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
