

Eukaryotic genomes from a global metagenomic data set illuminate trophic modes and biogeography of ocean plankton
- PMID: 37947402
- PMCID: PMC10746220
- DOI: 10.1128/mbio.01676-23
Eukaryotic genomes from a global metagenomic data set illuminate trophic modes and biogeography of ocean plankton
Abstract
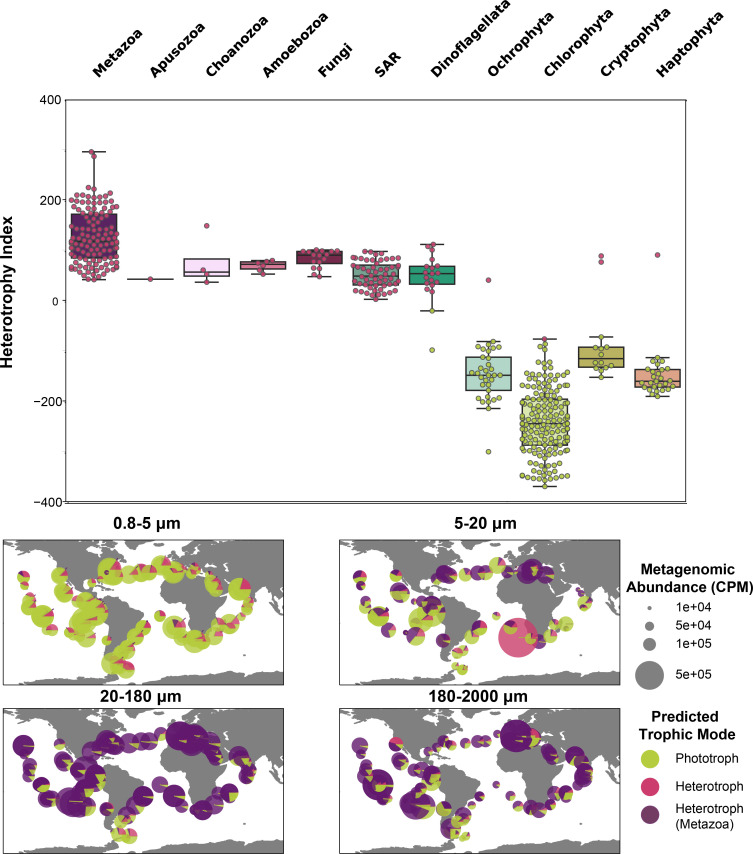
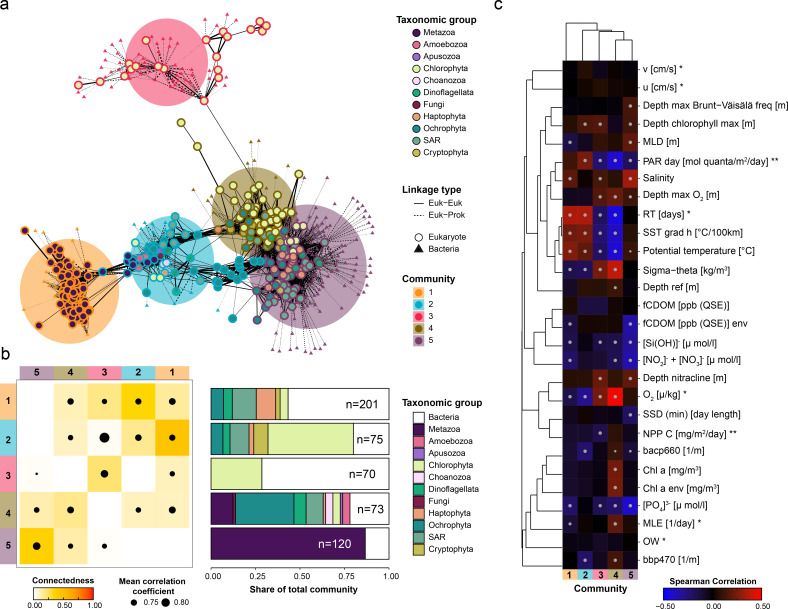
Single-celled eukaryotes play ecologically significant roles in the marine environment, yet fundamental questions about their biodiversity, ecological function, and interactions remain. Environmental sequencing enables researchers to document naturally occurring protistan communities, without culturing bias, yet metagenomic and metatranscriptomic sequencing approaches cannot separate individual species from communities. To more completely capture the genomic content of mixed protistan populations, we can create bins of sequences that represent the same organism (metagenome-assembled genomes [MAGs]). We developed the EukHeist pipeline, which automates the binning of population-level eukaryotic and prokaryotic genomes from metagenomic reads. We show exciting insight into what protistan communities are present and their trophic roles in the ocean. Scalable computational tools, like EukHeist, may accelerate the identification of meaningful genetic signatures from large data sets and complement researchers' efforts to leverage MAG databases for addressing ecological questions, resolving evolutionary relationships, and discovering potentially novel biodiversity.
Keywords: eukaryotic metagenome-assembled genomes; genomes; metagenomics; protists.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Mitra A, Flynn KJ, Burkholder JM, Berge T, Calbet A, Raven JA, Granéli E, Glibert PM, Hansen PJ, Stoecker DK, Thingstad F, Tillmann U, Våge S, Wilken S, Zubkov MV. 2014. The role of mixotrophic protists in the biological carbon pump. Biogeosciences 11:995–1005. doi:10.5194/bg-11-995-2014 - DOI
-
- Caron DA, Alexander H, Allen AE, Archibald JM, Armbrust EV, Bachy C, Bell CJ, Bharti A, Dyhrman ST, Guida SM, Heidelberg KB, Kaye JZ, Metzner J, Smith SR, Worden AZ. 2017. Probing the evolution, ecology and physiology of marine protists using transcriptomics. Nat Rev Microbiol 15:6–20. doi:10.1038/nrmicro.2016.160 - DOI - PubMed
-
- Caron DA, Countway PD. 2009. Hypotheses on the role of the protistan rare biosphere in a changing world. Aquat Microb Ecol 57:227–238. doi:10.3354/ame01352 - DOI
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
