

Telomeres as hotspots for innate immunity and inflammation
- PMID: 37951043
- PMCID: PMC10842095
- DOI: 10.1016/j.dnarep.2023.103591
Telomeres as hotspots for innate immunity and inflammation
Abstract
Aging is marked by the gradual accumulation of deleterious changes that disrupt organ function, creating an altered physiological state that is permissive for the onset of prevalent human diseases. While the exact mechanisms governing aging remain a subject of ongoing research, there are several cellular and molecular hallmarks that contribute to this biological process. This review focuses on two factors, namely telomere dysfunction and inflammation, which have emerged as crucial contributors to the aging process. We aim to discuss the mechanistic connections between these two distinct hallmarks and provide compelling evidence highlighting the loss of telomere protection as a driver of pro-inflammatory states associated with aging. By reevaluating the interplay between telomeres, innate immunity, and inflammation, we present novel perspectives on the etiology of aging and its associated diseases.
Keywords: Aging; Cancer; Genome stability; Inflammation; Innate immunity; Telomeres.
Copyright © 2023 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors of the review entitled ‘Telomeres as hotspots for innate immunity and inflammation‘ declare no conflict of interest.
Figures
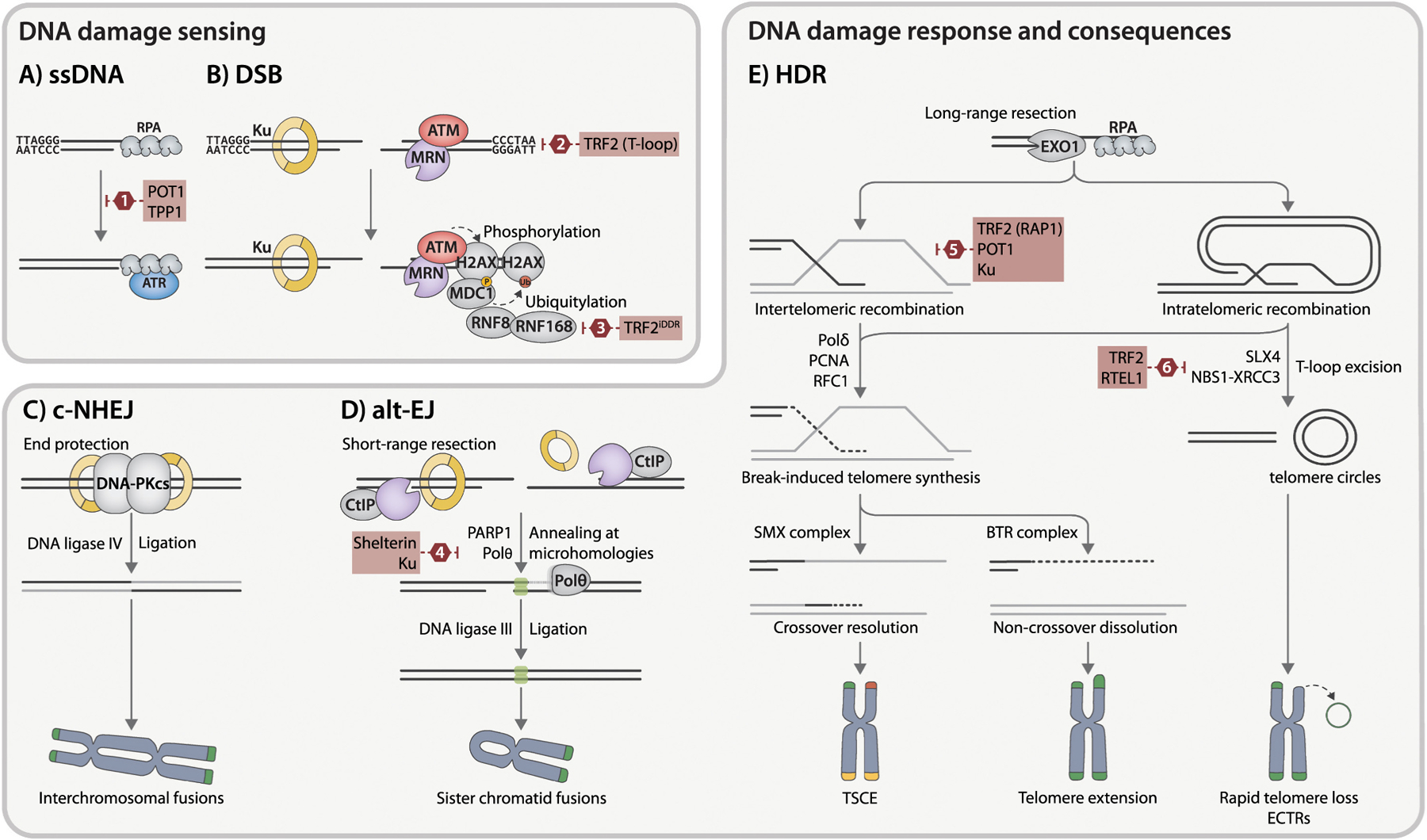
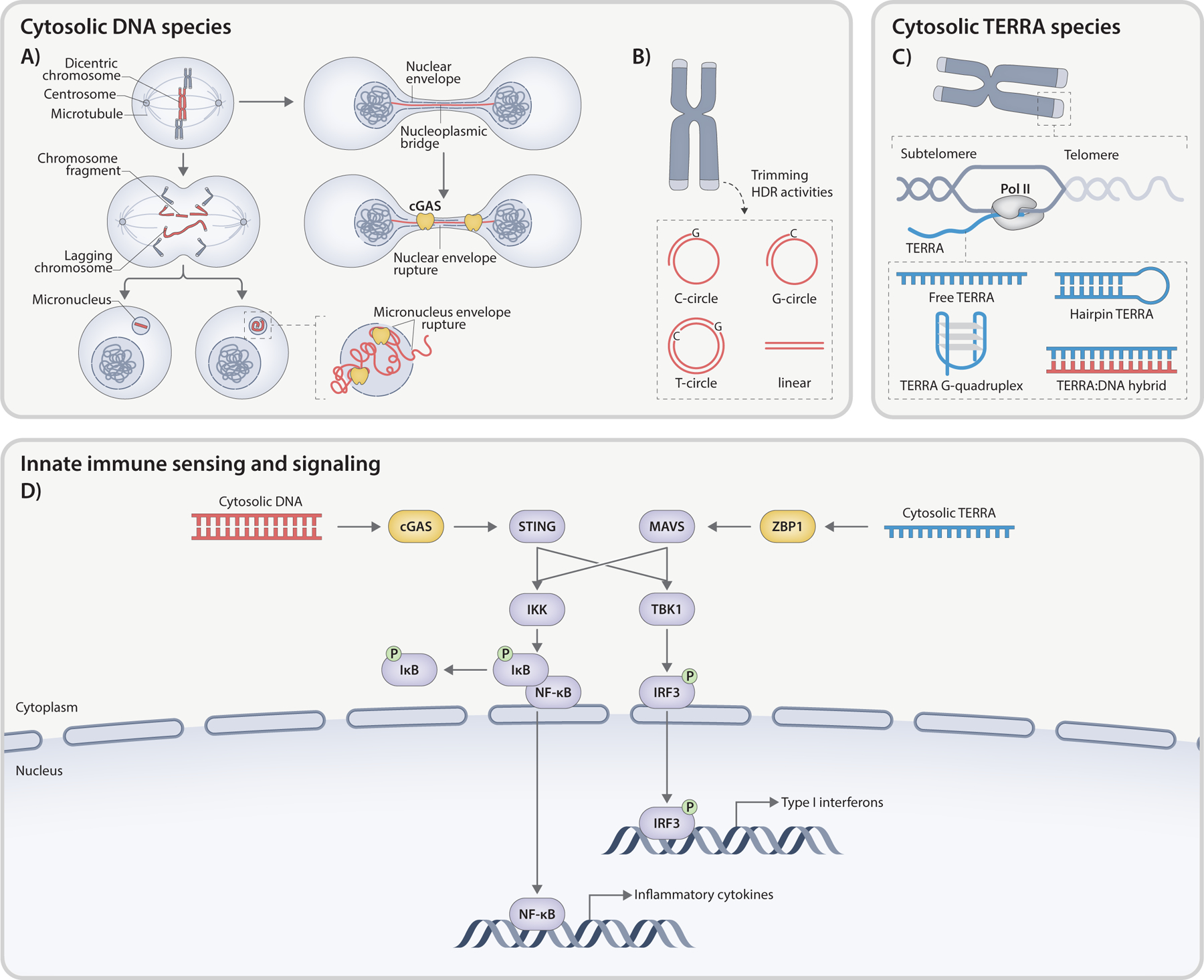
References
-
- de Lange T, Shelterin: the protein complex that shapes and safeguards human telomeres, Genes Dev 19 (2005) 2100–2110. - PubMed
-
- de Lange T, Shelterin-mediated telomere protection, Annu. Rev. Genet 52 (2018) 223–247. - PubMed
-
- Palm W, de Lange T, How shelterin protects mammalian telomeres, Annu. Rev. Genet 42 (2008) 301–334. - PubMed
-
- Sfeir AJ, Chai W, Shay JW, Wright WE, Telomere-end processing the terminal nucleotides of human chromosomes, Mol. Cell 18 (2005) 131–138. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
